
Profiling Flight Performance of Young Racing Pigeons (Columba Livia) in Training
Abstract
Two groups of young pigeons were trained and flight parameters profiled over a flight path of approximately 90 km and a number of measures were developed to analyse their flight performances. An aerodynamic model of avian flight was fitted to the data to assess its value in predicting flight performances. Both groups, although trained and flown independent of each other, had very similar profiles through time. Race speeds home peaked between weeks 5 - 8 of training and averaged 15.0 (2.0) m/s for Group 1 and 15.6 (2.7) m/s for Group 2. Flight performance in early stages of training in this study seemed to be affected by previous health conditions but once training progressed beyond 5 weeks, young pigeons appeared to fly at a constant preferred speed.
Article Information
- Received
- Accepted
- Published
Academic Editor: Mohammed Elmetwally, Assistant Prof of Theriogenogy, USA.
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2018 F.T. Scullion, et al
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Corresponding author: F. T. Scullion, Royal (Dick) School of Veterinary Studies, University of Edinburgh, 16 Upper Cranlome Road, Ballygawley, Co Tyrone, Northern Ireland, BT70 2HY —
Competing Interests
The authors have declared that no competing interests exist.
Funding
No specific funding statement was provided by the authors.
Data Availability
No data-availability statement was provided by the authors.
Acknowledgements
Thanks to all the pigeon fanciers who helped with the donation of birds, to Frederic Basch (MegaSystem, Deil Electronics BV) for the donation of the electronic timer and leg rings used in this study and to Leslie Madden for help with installation of the timer. Thanks to Professors Meredith and Pennycuick for encouragement. Thanks is also due to Paul Burr and the staff at Biobest for performing the PCR testing and to the staff at AFBI Omagh for microbiology laboratory work.
Citation:
Introduction
Homing pigeons (Columba livia) have an exceptional ability to navigate over long distances and if released at unfamiliar locations, instinctively and immediately endeavour to return to their home lofts. This homing phenomenon is the basis for the sport of pigeon racing - a popular hobby throughout the world1. Despite the homing pigeon having been the subject of much study in relation to avian navigation 2, 3, 4, 5, 6, 7, 8, 9, 10, avian flight mechanics 11, 12, 13, 14, 15, 16, 17, 18, 19 and basic physiological 20, 21, nutritional 22, 23and anatomical 24, 25, 26, 27, 28, 29 information, there are surprisingly few scientific studies on factors that affect racing performance.
In early works where pigeon homing behaviour was used to study avian navigation it became clear that homing ability was likely to be governed by a number of independent factors and doubts were expressed that observations of homing would be useful in determining the basis of avian navigation 3, 4, 5. An attempt was made to distinguish between orientation, describing the ability of a pigeon to head in the direction of home from a release site, and homing ability, describing how well, in terms of flight speed or percentage return rate, pigeons managed to get home 3, 4, 5, 10. Emphasis in navigation studies concentrated on time and direction of initial orientation and little or no effort was made to investigate specific factors that might affect flight speed in relation to racing pigeons' homing ability 10.
Diagram 1 is a schematic that outlines some of the major factors, from empirical deduction, that could have an effect on racing performance in pigeons.
Diagram 1. Major factors that could affect racing performance in pigeons.
Download figure
By way of example, in veterinary science, poor race performance has been listed as a clinical sign in association with adenovirus infection, circovirus infection, paramyxovirus infection, coccidiosis, chlamydophilosis and helminth burdens but there is little scientific evidence presented within these papers to support these claims 30, 31, 32, 33, 34, 35.
While empirical evidence derived from areas of scientific study, including avian medicine and pathology, navigation, flight mechanics, sport science, anatomy, physiology, biochemistry and nutrition may improve knowledge of, and suggest factors that may affect racing performance in homing pigeons, pigeon racing would benefit from a more scientific analysis of these factors.
It has been argued that scientific experimental investigation of sports can use methods that are so far removed from the actual sport as to be worthless. In order to be valid, studies on sport performance need to involve sport competitions or to simulate, as closely as possible, what happens in real events with athletes sampled from the population of interest 36. Currently however, routine racing protocols and performance measurements are not amenable to repeatable studies in racing pigeons and these require appropriate adaptation before progress can be made.
Matthews’ seminal works on avian navigation suggested speed improvements of homing pigeons during training may be due to actual differences in flight speed, but in the main will reflect the extent of deviation from the straight track home, or of halts on the way 3. However, the relationship between these aspects of flight performance in racing birds needs to be further defined and investigated.
In this study two groups of birds were trained for 10 weeks over increasing distances. A number of measures of speed including ground speeds and air speeds were analysed and profiled. An aerodynamic model was fitted to the results to assess its value in predicting minimum and maximum speeds 37 and a number of measures were created as an adjunct to investigating racing pigeon performance.
The first aim of this study was to describe the performance profile of young racing pigeons in training. The second aim was to evaluate the usefulness of new measurement techniques and aerodynamic theories in determining how performance characteristics in young racing pigeons change throughout training.
Materials and Methods
An observational study was carried out on two groups of pigeons (Group 1 and Group 2) trained to fly along an 87.02 km path for a period of 10 weeks. The husbandry, management and training methods as described in this paper are those normally used by pigeon fanciers in the sport of pigeon racing. Health assessments and treatments were carried out as a veterinary surgeon, under the Veterinary Surgeon’s Act 1966.
Loft Construction
A 10.0 x 3.3 x 2.3 m metal framed, south facing pigeon house built on concrete block stilts was constructed using corrugated iron cladding and internal timber lining. 2.0 x 2.3 x 2.3 m sections were separated off as individual lofts for breeding stock and young pigeons. Lofts used in this study were randomly chosen. Large wire mesh windows, that could incorporate racing pigeon traps, were placed centrally on the south facing wall of each loft. Floor ventilation slots were inserted in the access corridor so that ventilation in the bird section could be controlled. The north wall of the young pigeon lofts was covered with box perches sufficient in number to allow all birds to have their own perch and view the outside of the loft from their perches.
Acquisition and Management of Pigeons
Young racing pigeons are generally hatched as early as possible in the year and some fanciers occasionally allow racing birds to pair up again, lay eggs and hatch further young as a method of encouraging them to race home in the adult race season. Some of the latter young birds are in excess of requirements for the young bird race season. Timing for the adult race season means they are generally hatched in 'rounds' in early March and in mid-April. A request was put to fanciers for donations of such birds for this study. Allocated birds were between 6 and 8 weeks old. It has been shown that breed has an effect on homing ability 3 and although all the birds donated were from racing pigeon stock, stratified random sampling was carried out so that birds from each loft of origin were evenly distributed throughout the study groups. Birds were randomly allocated, within the loft of origin strata, to each group by drawing numbers blindly from a hat and groups allocated to individual lofts. Initially, birds were allowed a two week settling in period before training commenced. Water was provided ad libitum. A standard seed mix, consisting of yellow maize, wheat, mixed peas, milo, dari, striped sunflower seeds, kardi seeds, Australian peas and buckwheat, was provided once each day, at a rate of 50g per bird 38.
Training
Training followed the techniques used by pigeon fanciers, with food as the perceived motivation to home 2, 3. Feeding was at 17:00 each day. Birds quickly learned when their feed was due. The first training phase(pre-training) which lasted approximately two weeks, involved getting birds accustomed to exiting and entering the loft. This was accomplished by placing a temporary aviary outside the opened loft window so that birds had free access to a small safe area outside which allowed them to perch and view the surrounding countryside. A trap system was then placed in the window opening. The one way exit door could be left open to allow the pigeons out to the aviary and they could enter the loft again via the entrance trap door at feeding time. Once all the birds were comfortable in exiting and entering the loft, they were allowed to exercise around the loft on a daily basis. Initially, birds spent a lot of time perching on the loft roof but gradually they ventured away for flights of up to 1 hour duration. At this time basket training was initiated when birds were placed in a carrier basket (basketed) and taken up to a few hundred metres from the loft and released (tossed). About 3 weeks into their training the daily routine would involve basketing birds, taking them away from the loft for a short distance, tossing them and allowing them to fly back to the loft where they would be called in for food. In the final phase (actual training), when studies were begun, the birds were trained on a continuous line of the chosen trial flight path (Table 1). At this stage birds were either having free flight around the loft or, weather permitting, trained daily on the trial flight path by release at increasing distances.
Table 1. Release sites, beeline distance from release site to the loft, map co-ordinates of various sites and bearing to the loft from the release site| Release Site | Beeline Distance (km) | Latitude | Longitude | Bearing(degrees) |
| Loft | 0 | 54.508147 | -6.983166 | |
| A | 6.935 | 54.4701 | -6.898084 | 307 |
| B | 7.27 | 54.484037 | -6.878501 | 291 |
| C | 13.39 | 54.500502 | -6.776247 | 273 |
| D | 13.81 | 54.477772 | -6.775817 | 256 |
| E | 21.29 | 54.496698 | -6.65404 | 273 |
| F | 23.9 | 54.481591 | -6.615888 | 277 |
| G | 39.35 | 54.480974 | -6.375605 | 274 |
| H | 49.61 | 54.485388 | -6.215904 | 273 |
| I | 73.18 | 54.558763 | -5.852226 | 265 |
| J | 87.02 | 54.399598 | -5.650063 | 277 |
Release points were chosen to give a clear all round view, away from road traffic and overhead electric wires with no trees or buildings near enough to obstruct take off or block the view of the observer monitoring initial progress. There is a strong tendency for pigeons to join up with any other group of pigeons, so birds were not released on race days and the training path chosen was on a different line from the majority of actual pigeon races. Each group of birds was released into the air from their carrier basket held aloft by the liberator in a randomly chosen direction. The next group of birds were not released for 20 minutes or until, over short distances, word was received that the preceding group had actually arrived home. In this way, the results from individual groups of birds were independent of each other.
The extent of the training path used was approximately 90 km east of the home loft. Releases occurred at various intervals along this flight line over the period of the study (Table 1). Birds trained to fly the full 90 km were kept fit with shorter training flights or free flights around the loft on alternate days.
Aerodynamics
A computer programme, Flight 1.24, was used to calculate aerodynamic parameters of flight. Full details of the theories and the workings of the Flight programme used in this study can be found in Pennycuick 37.
The Flight programme generates a mechanical power curve with air speed along the x axis (see below for definition of air speed). This shows the rate at which the flight muscles must do mechanical work in steady, level flight as a function of air speed. This programme uses aerodynamic theories to predict a bird’s flight performance and derives power usage at different air speeds in forward flapping flight. The curve of power versus speed for horizontal flight has a minimum power speed (Vmps) where the power consumption per unit of time is lowest 11. Birds are not expected to fly slower than Vmps for any extended periods since it would result in flight instability. In wind tunnel experiments (where birds are in stationary flight at the speed of the wind), birds tend to stop flying within a short period of time when wind speeds are below Vmps 39, 40, 41. There is also a maximum range speed (Vmrs) at which a bird can fly the greatest distance using the minimum amount of energy 11. Biological scientists associate the Vmrs with endurance performances such as bird migration 42.
Air speed varies with air density which is related to altitude. In this study the furthest release point was situated at sea level and the loft is on a hill at approximately 204 metres. The actual height at which the pigeons flew was unknown so an average altitude of 102 metres was used in the Flight programme. Apart from this and the input data mentioned below, the rest of the variables were set at the defaults suggested in the Flight programme.
Diagram 2 illustrates the wing span and wing area measurements, which along with the mass of the birds, are entered into the programme. The wing measurements were calculated as directed in the programme, by holding a bird on its back on a sheet of paper and scribing the necessary outline, once for each bird prior to commencement of training. Wing span was measured with a ruler and wing area by using an acetate grid and counting 1.0 cm2 squares. The mass of the birds was recorded and entered prior to each flight.
Diagram 2. Illustrating the measurement of wing span and wing area.
Download figure
Other variables derived from the Flight programme output included an individual’s Flight Performance Ratio (FPR) expressed as a percentage measure of the air speed in flight in relation to the Vmps and Vmrs.
Speeds
The speed of a racing pigeon can be defined relative to either the ground or the air. Both speeds are affected by wind speed and direction. The relationship between ground speed, air speed and wind speed are related by vector mathematics.
Ground speed
An individual bird’s ground speed is calculated by dividing the beeline distance between release site and loft in metres, by flight time in seconds. In pigeon racing the race speed is the ground speed. The observer noted release times when the birds were freed from the carrier basket. Return times were recorded using an electronic timer system (MegaSystem, Deil Electronics BV) for individual birds arriving back in large groups. The time recording antenna was positioned so that the bird’s fitted leg ring would be read as the bird made its way from the landing platform on the outside of the trap door to the reception area inside the loft. Flight return times were downloaded from the timer clock for all birds after each flight.
Air speed
Air speed is the speed of the bird relative to the air and is used in aerodynamic models of bird flight 11. Air speed was calculated using wind triangle trigonometry and vector calculations similar to those used by pilots in navigation 43, 44, 45. By convention, wind direction is recorded as the direction from where the wind is blowing. However, the direction to where the wind is blowing is used in the air speed calculations. The ground speed and bearing between release site and home and the wind speed and direction vectors are parameters required to calculate air speed.
Wind Speed and Direction
The most difficult variables to measure accurately in relation to bird flight are the wind speed and direction because they vary substantially between sites and at different times 41. Due to budgetary constraints and limited time available it was beyond the scope of this study to measure wind speed and direction at more than two locations.
The wind speed was measured, simultaneously, three times within one minute at the loft and the release site using a digital anemometer (Holdpeak HP-816A, Zhuhai Jida Huapu Instrument Co., Ltd., Guangdong, China). Wind speed was measured at the release point, approximately 3 m from the ground, by standing on an isolated platform and holding the instrument at arm's length in the air and on the loft roof, approximately 5 m from the ground, by standing on the centre of the roof of the loft and holding the instrument at arm's length in the air. Wind direction was measured three times using a hand held compass (Silva Classic Compass, Sports Warehouse, Edinburgh, UK) with a fine thread attached. At a single location, the wind speed measurements taken deviated by less than 25% and the wind direction varied by less than 10 degrees. Average wind speed and direction were calculated from loft and release site figures for each training flight using vector mathematics. Birds in this study were estimated to fly at an average height of approximately 102 m for calculations in the Flight programme. Measured wind speeds were corrected to estimates of wind speed at 102 m using techniques employed by the wind turbine industry 46. However, vertical wind components were not measured by the techniques available and could not be included in the calculations.
Wind Vector
The ground speed and the air speed of a bird are considered to be equivalent in zero wind conditions 47,48. The wind vector Vw can be calculated as where Vg is the scalar vector quantity of the ground speed and Va is the scalar vector quantity of the air speed. In a plot of Va against Vw the regression can be calculated where a is the intersection with the Y axis and b is the slope of the regression line of the graph and both a and b are constants. Then it follows and when Vw is zero
In other studies a dimensionless speed measurement has been used in order to generalise between different species 47, 48. It is useful to know the speed at zero wind conditions so that direct comparisons can be made between birds at different times as it is a dimensionless measurement. In this study the FPR is a further dimensionless measurement. The wind vector can be standardised by dividing by the Vmps. When FPR is plotted against the standardised wind vector, the FPR in zero wind conditions can be found. The FPR of birds in zero wind conditions can also be used to determine the air speed in zero wind conditions by back calculation from the average Vmps and Vmrs of the birds under study.
Preventative Medicine
A number of techniques, similar to those used by fanciers, were put in place in an attempt to avoid any confounding effects of the most common disease causing organisms on race performance.
Veterinary clinical examinations similar to the method described in Coles 49 were carried out on each bird throughout the study. Laboratory diagnostic tests were performed on each group of birds prior to commencement of training (pre-training health tests).
Briefly, individual birds were weighed and on handling, the birds' mouths, nares, ears and eyes were checked for visible lesions. Heads and necks were palpated and wings opened to inspect feathering on the dorsal and ventral sides. Shoulder, elbow and carpal joints were palpated, extended and flexed and bones and soft tissues of the wings palpated. The body including pectoral muscles and keel and abdomen were palpated and the skin covering the breast inspected. The hip, stifle, tarsometatarsal and toe joints were palpated, extended and flexed and bones and soft tissues of the leg palpated. Foot pads were inspected. The heart, air sacs and lung fields were auscultated. Two individual pigeons were detected to have been injured by hawks during this study. These birds were rested and given appropriate treatment until deemed fit to fly again. Natural moulting of feathers was detected in some of the birds in week 10 which resulted in the curtailment of the studies. Prior to commencement of training the birds were affected by a "Young bird sickness" clinical outbreak associated with a secondary viral infection 50.
All the birds were vaccinated prior to the commencement of training against Paramyxovirus (Colombovac® PMV, Pfizer Ltd.) and groups of birds were checked by means of pre-training health tests, for the presence or absence of pathogens that are readily treated and commonly found in young pigeons. Droppings collected over a 5 day period were submitted to a commercial laboratory (Biobest, Edinburgh) and tested for the presence of Chlamydophila using standard PCR protocols 51. Throat swabs were taken from three birds and fresh samples of droppings were examined using techniques described in Scullion and Scullion 52. Briefly, throat swabs were stored in 5 ml of trichomonas medium (E&O Laboratories Ltd, Edinburgh) and examined within 8 hours. The three throat swabs were smeared on a single glass slide, covered with a coverslip and examined under the light microscope for the presence of trichomonads, Candida spp. and bacteria. Droppings were stored in a sealed plastic bag until they were examined within 8 hours. A small sample of faeces was taken and mixed in a drop of tap water on a glass slide, covered with a coverslip and examined under a light microscope. These smears were examined for the presence of Coccidia oocysts and Ascarid and Capillaria worm eggs, Candida and bacteria. A further sample of droppings was submitted to a commercial laboratory (AFBI Omagh) for microbiological culture and sensitivity testing 53.
In the "Young bird sickness" clinical outbreak, sick birds were passing sticky green droppings and were fluffed up and thin. Four birds died. The secondary virus was provisionally diagnosed as an Adenovirus using cytology, due to the finding of basophilic intranuclear inclusion bodies surrounded by a halo, in hepatocytes 31, 54, 55. All birds were confined to the loft and treatments were applied for secondary infections of E.coli, Canker, Coccidia, Capillaria and Candida using treatments as described in Scullion and Scullion 56. The clinical signs abated after approximately three weeks, as determined by veterinary clinical examinations described above.
Other measurements
Prior to training birds were randomly assigned to each group and each loft. There were 23 and 22 birds in Groups 1 and 2 respectively.
Slow returns home were recorded in each race. Slow returns were defined as birds taking at least twice as long to return home as the fastest birds in their group. Slow returns were included in calculations of average speeds for the groups unless they returned more than six hours after their compatriots when their results were excluded on that occasion.
Vmps and Vmrs were calculated for each individual bird flight for each training session. Weekly average Vmps and Vmrs were calculated for each group. Average combined Group 1 and Group 2 potential scalar vector quantities for ground speeds (m/s) each week (Vg) at aerodynamic air speed delimiters (Vmps and Vmrs) were calculated by adding Vw to Vmps and Vmrs 47, 48. Using these ground speeds, theoretical distances (D) were derived from
Vg = D/T
where T was the actual time recorded for the flight (seconds). Flight efficiencies were then calculated by dividing the beeline distance (m) by D (m) to assess how the birds' flight may be affected by air speeds of Vmps or Vmrs.
FPR versus wind vector/Vmps regressions were plotted using data from the fourth week onward and the regressions were used to calculate zero wind FPRs and speed at zero wind for Groups 1 and 2 respectively.
Results
The two groups of birds were each given 43 tosses over a 10 week period of actual training over a total beeline distance of 1027 km (Figure 1). Since it is unknown if the birds flew the accurate beeline distances on each flight the distances flown are expressed as minimum distances. Together the birds spent over 61 hours away from the loft during training. Birds were flying the maximum distance on the training path by week 5 of training.
Figure 1. Number of training tosses and minimum distances flown per week by each group. Flocks of pigeons generally fly as single entities.
Download figure
Data from the two groups were compared for major differences. Flocks of pigeons generally fly as single entities. This excludes the ability to analyse the differences between the two groups by statistical means 57, 58. For the majority of training tosses, birds in each group acted similarly with only seconds of difference in training flight times on occasion. This was despite there being sufficient time between releases to prevent the later group from being influenced by the behaviour of the first group. A few tosses resulted in flight times with major differences in return times of the two groups or when an individual or two were noticeably slower in returning to the loft. These flights were either associated with poor weather conditions, such as heavy rainfall occurring when the birds were en route from the release site, or when there was evidence of hawk strike. It was felt either the birds had stopped to roost during the rain or were driven off their flight path by the action of predators.
Slow return and loss data was assessed in relation to the two lofts combined. At the start of the study there were 45 young birds. 4 birds died during the sickness episode before pre-training commenced. By the time of actual training there were 18 birds in Group 1 and 18 birds in Group 2. At the end of the study there were 20 young birds left with 11 birds in Group 1 and 9 birds in Group 2. In terms of individual flights of birds over the actual training period there were a total of 951 individual flights and 21(2%) birds were lost in total during training. Some losses (5 birds) occurred during the pre-training phase in the few weeks of free flights around the home loft before the actual training sessions commenced (Figure 2). The number of birds that were lost during actual training was highest in the first four weeks (12 birds in the first four weeks). One of the lost birds was later reported to have crossed the Irish Sea and entered a loft in the North of England.
Figure 2. This shows the fortnightly number of individual bird flights where birds returned home late, or were lost, over the 10 week training period in both lofts. Slow returns were defined as birds taking at least twice as long to return home as the fastest birds in their group.
Download figure
Slow returns were not measured in the pre-training phase. Of the 951 individual bird flights in actual training, 45 (5 %) were classed as slow returns. The number of birds that returned home slow was highest in the first two weeks of actual training.
Mean ground speeds (SD) achieved over the 10 weeks varied but both groups followed a similar pattern rising to a peak between weeks 5 - 8 of 15.0 (2.0) m/s for Group 1 and 15.6 (2.7) m/s for Group 2 (Figure 3 and Figure 4). The slowest ground speeds occurred during the initial three weeks of training flights of the young birds.
Figure 3. Weekly ground speeds of Group 1. Since the actual distance flown may be more than the beeline distance then the accuracy of the resultant ground speeds is unknown.
Download figure
Figure 4. Weekly ground speeds of Group 2. Since the actual distance flown may be more than the beeline distance then the accuracy of the resultant ground speeds is unknown.
Download figure
Both groups showed similar air speed patterns during training. Air speeds for each group only exceeded the groups’ mean Vmps after the first three weeks of training (Figure 5 and Figure 6). Both groups of birds had air speeds within the mean Vmps and Vmrs of the respective groups between weeks 4 and 9 inclusively. Air speeds in the final week fell below Vmps in both groups. The average air speed(SD) for Group 1 between weeks 4 and 9 was 16.9(1.8) m/s and that for Group 2 was 17.4(2.0) m/s when air speeds fell within the Vmps and Vmrs speeds predicted by the aerodynamic model.
Figure 5. Weekly air speeds compared with the aerodynamic delimiters of Group 1 birds.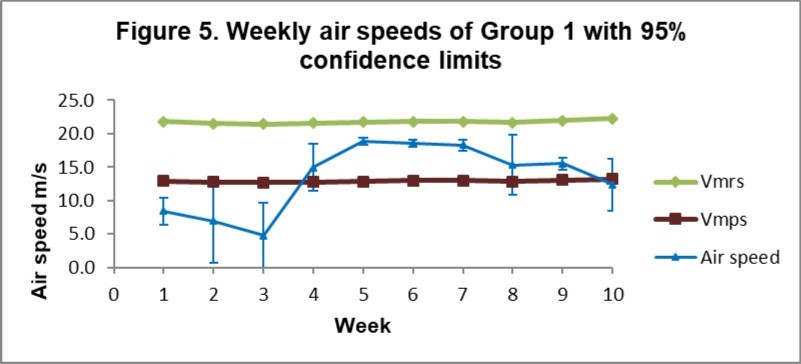
Download figure
Figure 6. Weekly air speeds compared with the aerodynamic delimiters of Group 2 birds.
Download figure
Figure 7 illustrates the FPRs for Groups 1 and 2. This measurement makes it easier to compare performance of each group as it standardises their aerodynamic abilities. The pattern of performance is similar throughout the training sessions with Group 2 slightly outperforming Group 1 overall, having an average performance ratio of 13% compared to 6%. In weeks 1-3 and 10 the FPR suggests flight speed is such that the birds are either resting up or the distances flown are much more than the expected beeline distances.
Figure 7. Weekly flight performance ratios (FPR) of Groups 1 and 2. This measurement makes it easier to compare performance of each group as it standardises their aerodynamic abilities. A negative FPR occurs when the air speeds are less than Vmps when according to aerodynamic theory the bird is unlikely to fly. Air speeds can appear to be low when actual flight distances are increased well beyond beeline distances.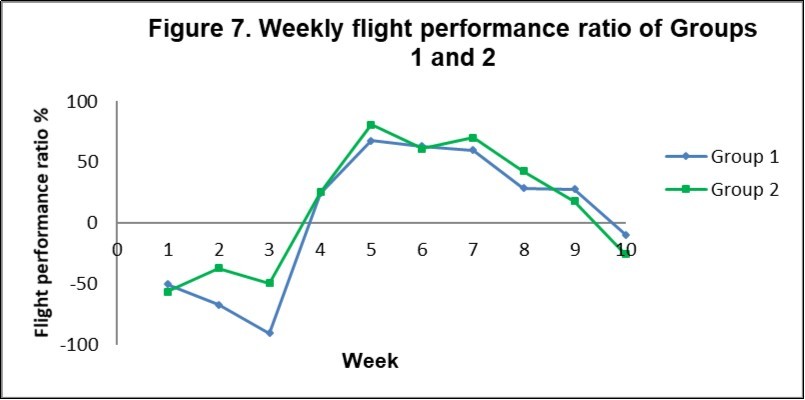
Download figure
Figure 8 illustrates the flight efficiency of the birds each week if it is postulated that they fly at Vmps or Vmrs. During weeks 5 – 7, if the birds fly at Vmrs they have a flight efficiency that is near 1.0 which would mean they are flying straight home.
Figure 8. Flight efficiency of the birds each week if it is postulated that they fly at Vmps or Vmrs. Flight efficiency can be calculated as the ratio of the beeline distance/ total distance flown. Perfect flight efficiency, straight along a beeline, between release site and home loft is 1.0. Flight efficiency cannot be greater than 1.0.
Download figure
Figure 9 and Figure 10 show the FPR v wind vector/Vmps for Groups 1 and 2 respectively. For Group 1, the zero wind vector has a ratio of 3.6%, which equates to a speed at zero wind of 13.3 m/s. For Group 2, the zero wind vector has a ratio of 9.4%, which equates to a speed at zero wind of 14.0 m/s.
Figure 9. Flight performance ratio versus wind vector/Vmps for Group 1 birds.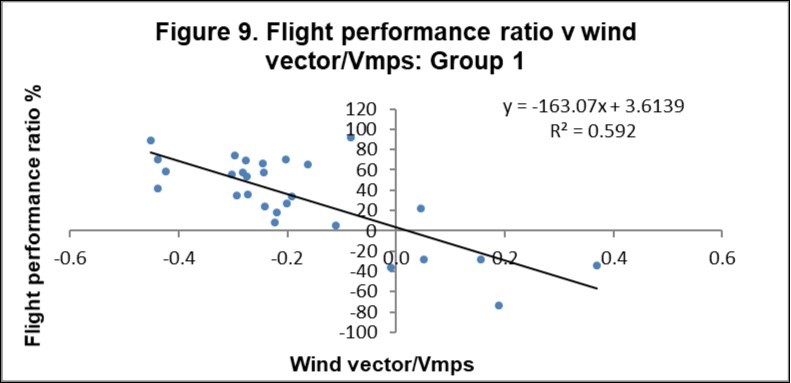
Download figure
Figure 10. Flight performance ratio versus wind vector/Vmps for Group 2 birds.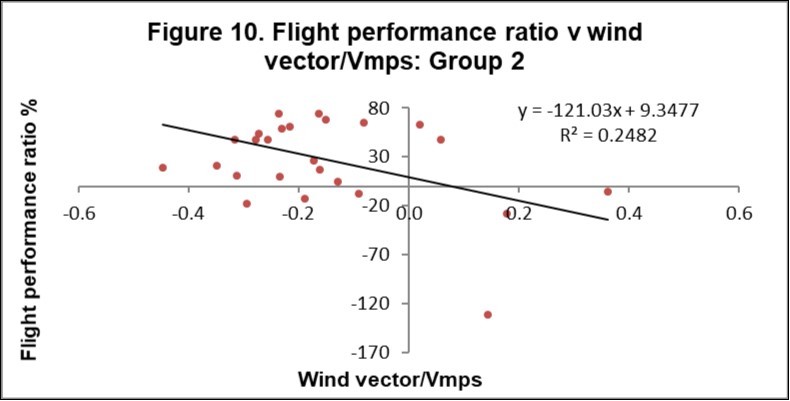
Download figure
The birds began to moult in the 10th training week and a few birds were noted to have lost some primary wing feathers and some tail feathers which are vital for flight and this is reflected in the decreasing performance. Training was halted at this stage to prevent loss of birds.
Discussion
The calculated distance between the loft and the furthest release site of 87.02 km is comparable to the first race distances that young birds fly during the racing season. The birds were held, managed and trained with methods used by pigeon fanciers. The data collected from these racing pigeons represented two independent groups and the trends illustrated serve as a performance profile for young racing pigeons during a ten week training period. The performance profile from each group was relatively similar, with releases on the same day showing similar speeds of return. By all measures used, the flight performance improved in both groups over the first four weeks, peaked between the fifth and eighth weeks after which there was a drop in performance associated with the moulting of feathers in the birds.
In this study, excluding losses due to sickness prior to commencement of training, losses of young pigeons around the loft and during training were 5/41 (12%) and 16/41 (39%) respectively. This is higher that the average losses of 8% and 16% per loft, around the loft and during training respectively, in a single season in 28 pigeon lofts in Scotland 59. The young birds in this study had apparently recovered from “Young bird sickness”, by the time they were being flown in pre-training and actual training but undetected subclinical effects may have left the pigeons less able to orientate and navigate in flight and more susceptible to loss and/or predation 50.
Although early studies on navigation in pigeons suggested an innate aspect to homing behaviour 4, 5, 60, improvements occurred after a short course of training flights from 10 - 25 miles (approx. 15 - 40 km) 3, 61, 62 or within 3 to 6 releases from the same site 10, 63. In this study, improvement appeared strikingly delayed compared to the navigation studies mentioned above. Despite a lack of knowledge of the accuracy of ground speeds, this study showed that performance improvement was only recognised in the fourth week of training when birds had flown approximately 300 km over 16 tosses. After the fourth week changes in slow returns and losses, ground speeds, air speeds, FPRs and potential flight efficiencies all suggested an improvement in the performance profile in both groups. By this stage both groups of birds were flying home at speeds above the minimum power speed predicted by aerodynamic theory. Vereecken et al. suggest that racing performances in pigeons can remain poor for several weeks after clinical recovery from adenovirus infection and speculate that young pigeons affected with adenovirus will gain less flight experience and thus fall behind their non-infected congeners 31. In this study an effect of the earlier adenovirus infection in the birds cannot be ruled out as a cause of the poor flight performance in the first four weeks of training. The explanation that the young birds have less flight experience than their congeners is less likely to be valid since all the birds had similar flight experience over the whole training period.
Pigeons trained to fly distances up to 65 km, followed by an aeroplane, using radio-tracking techniques, appeared to fly at a relatively steady air speed(SD) of 13.7(0.75) m/s 64. In this study, calculated air speeds, in the latter half of the training period, became less variable and were greater than Vmps speeds but less than Vmrs speeds predicted by aerodynamic theories. The average(SD) air speeds in groups 1 and 2 were 16.9 (1.8) m/s and 17.4 (2.0) m/s respectively between weeks 4 and 9. Terrestrial animals have a preferred speed within each type of gait, that is used more frequently than other speeds, and is associated with minimum energy cost per unit distance travelled 65. Pennycuick suggests that different species of birds' choice of flight speeds may be size related 41. It is possible that racing pigeons, when trained to home, pace their journey at a relatively steady flight speed although more accurate measurements of actual distances flown would be required to confirm this.
Vmrs is predicted, by aerodynamic theories, to be the speed at which a bird can fly the greatest distance at minimum energy costs and by definition is thought to be the preferred speed for endurance flight in birds, such as migratory flights 42, 66, 67, 68. Some postulate that birds fly at speeds that compromise between the minimum energy cost of flight, Vmps, and the minimum energy costs per distance, Vmrs 69. However, field studies on 32 bird species suggested that they migrate at speeds nearer Vmps than Vmrs 41. This study suggests that in the later training stages, the racing pigeons may have had an air speed that is near to the Vmrs and thus in well trained racing pigeons Vmrs may be the preferred flight speed. If racing pigeons do fly at a preferred speed and aerodynamic theories are correct then ideally birds should be trained to achieve a preferred speed of at least Vmrs. It would be of immense benefit to racing pigeon fanciers to have further studies that indicate methods of training and motivation that ensure racing pigeons use this optimum speed each time they race. However, this determination of Vmrs as the preferred speed is reliant on the validity of the aerodynamic theories as applied to the racing pigeon.
The Flight programme predicts a bird’s performance from a model based on accepted laws of physics, not from a statistical model of the kind more commonly used in biology. The programme is based on theories that represent the flight abilities of a bird, in much the same way that an aircraft design engineer represents the flight of an aircraft 37. Although much of the original theory was developed using trained pigeons in wind tunnels 11, the programme has proved to be robust in its use across a number of avian families in different scientific studies representing a large variation in bird size and shape 70, 71, 72, 73. There are limitations to the use of such a modelling method. In particular it excludes dynamic effects of vertical wind components and the effects of potential energy on birds in flight. However, despite its origins and limitations, to this author’s knowledge, this is the first use of the programme to investigate flight parameters in racing pigeons under conditions equivalent to those used by the fancy and it appears to be a valuable tool to assist in the investigation of racing pigeon flight.
It is difficult in this study to determine what exactly happened in the first few weeks of training since the distances flown are unknown and the birds may halt in their flight. To clarify these aspects, further studies are required that examine the actual routes of the birds, assess the validity of the aerodynamic theory and investigate the flight performance of young racing pigeons in the first four weeks of training in more detail.
While the times measured for training flights were accurate, actual distances flown were unknown, so theoretical aerodynamic air speed benchmarks Vmps and Vmrs, were used to predict distances flown which in turn were expressed as possible flight efficiencies. Perfect flight efficiency, straight along a beeline, between release site and home loft is 1. Any efficiency greater than 1 that results from such a calculation suggests the beeline distance is greater than the total distance flown. It is logical that the data shows that from the fourth week onwards, the birds have to be flying at a speed that is faster than the predicted Vmps, since the distance flown cannot be shorter than the beeline distance. Further, a steady air speed of Vmrs predicts the possible flight efficiencies trend toward 1 after the fourth week until the seventh week and the 95% confidence limits suggest that if actual speeds were much slower than this, efficiencies again would be greater than 1. The data and theory thus support the birds flying at a postulated constant speed near Vmrs from the fourth week onwards.
Calculations using a steady Vmrs speed throughout training, suggest that flight efficiency although initially low, rises toward perfect efficiency levelling off and remaining relatively constant for a 4 week period with a mean(SD) of 0.77(0.07) before gradually decreasing toward the end of the training period. This improvement in flight efficiency is similar to findings from early avian navigation studies, confirmed by recent studies using GPS tracking, where improvements in flight efficiencies resulting from a decrease in the distance of the actual flight path, occur as the number of flights from a release site increase 3, 10, 74, 75, 76. Thus fitting the data in this study to an aerodynamic model allows predictions that are consistent with findings from studies in avian navigation and suggest that young racing pigeons’ improvement in flight speed are to a large extent resultant from improvements in navigational abilities as was postulated by Matthews 3. The aerodynamic model also suggests that ground speed data becomes more reliable as training progresses, since distances flown veer towards beeline distances.
Generally the patterns of change in ground speeds, air speeds and FPRs in this study reflect their mathematical derivations involving their relationship with wind speed and direction. The main difficulty in their current usage is the reliance on calculations with unconfirmed distances flown by the birds and this is compounded by changing distances flown by improvements in navigation. Despite this there appears to be a relationship with measured speeds and predictions from aerodynamic theories which deserves further investigation.
This study introduces speed at zero wind as a further means of comparing flight performance between birds at different times. Dimensionless quantities are widely used in mathematics, physics, engineering and economics. The use of dimensionless numbers is also important in biomechanics since they allow comparisons between systems despite differences in medium or scale 77. Speed at zero wind is a standardised measurement calculated from a regression of dimensionless measurements that in theory enables such comparisons of flight performances at different times and under different conditions but its usefulness will need to be tested further.
The data in this study and the logical conclusions drawn from the use of aerodynamic theories suggests that young pigeons are capable of reaching maximum flight speeds by the fourth week of training and that major improvements in flight speed are related to improvements in navigation abilities. The aerodynamic theory used in this study was developed using pigeons in flight tunnels and remains to be validated for use in racing pigeon studies.
This study highlights a period in the first four weeks of training where young pigeons appear to perform poorly. Analysis of this period is limited due to the lack of knowledge of actual distances flown during this time and aerodynamic theory suggesting the birds are either flying extreme distances or not flying for part of the time away from the loft during this period. GPS technology is sometimes used by fanciers to monitor the routes their birds have flown 78 and is worthy of consideration in further studies of this early training phase. The birds used in this study were clinically healthy as determined by frequent veterinary checks and laboratory investigations for common pathogens that may affect flight performance. A subclinical effect of a recent adenovirus infection on flight performance cannot be ruled out during the early phase of training. The effect of health on flight performance is therefore worthy of further study.
Conclusions
1. Trained young racing pigeons appear to fly at a constant preferred speed.
2. Flight performance of young pigeons during early training stages may be affected by previous health conditions.
3. Aerodynamic theories of bird flight are of value in investigations of racing pigeon flight performance but further validation studies are required.
4. A number of measurements including ground speed, air speed, Flight Performance Ratio, speed at zero wind and flight efficiency have been defined that are useful for investigation of racing pigeon flight performance.
References
- 1.Doneley B. (2006) Pigeon medicine and surgery. Proceedings North American Veterinary Conference 20, 1525-1530.
- 3.GVT Matthews. (1951) The experimental investigation of navigation in homing pigeons. , J. Exp. Biol 28, 508-536.
- 4.GVT Matthews.(1953a). The orientation of untrained pigeons : A dichotomy in the homing process. , J. Exp. Biol 30, 268-276.
- 7.Keeton W T. (1970) Comparative orientational and homing performances of single pigeons and small flocks. , Auk 87, 797-799.
- 8.Papi F. (1982) Olfaction and homing in pigeons: Ten years of experiments. In Avian Navigation. Papi F and Wallraff HG (eds) , Berlin. Heidelberg. New York: 151-159.
- 9.Papi F. (1986) Pigeon navigation: solved problems and open questions. , Monit. Zool. Ital. (NS) 20, 471-517.
- 10.Wallraff H G. (2005) Avian Navigation: Pigeon Homing as a Paradigm. , Berlin, Heidelberg, Germany
- 11.Pennycuick C J. (1968) Power requirements for horizontal flight in the pigeon Columba livia. , J. Exp. Biol 49, 527-555.
- 13.Pennycuick C J. (1975) Mechanics of flight. In Farner DS and King JR (eds). Avian Biology. (5) , New York: 1-75.
- 15.Tucker V A. (1973) Bird metabolism during flight: Evaluation of a theory. , J. Exp. Biol 58, 689-709.
- 20.Butler P J, West N H, Jones D R. (1977) Respiratory and cardiovascular responses of the pigeon to sustained level flight in a wind-tunnel. , J. Exp. Biol 7, 7-26.
- 21.Biesel W, Nachtigall W. (1987) Pigeon flight in a wind tunnel. IV. Thermoregulation and water homeostasis. , Journal of Comparative Physiology B: Biochemical, Systemic and Environmental Physiology 157(1), 117-128.
- 22.Vogel L, Gerlach H, Löffler M. (1994) Columbiformes. In Ritchie BW, Harrison GJ and Harrison LR (eds). Avian Medicine: Principles and Application. Wingers Publishing Inc , Lake Worth, Florida: 1200-1217.
- 23.Sales J, GPJ Janssens. (2003) Nutrition of the domestic pigeon. , World’s Poultry Science Journal 59(6), 221-232.
- 24. (1988) A functional analysis of the primary upstroke and downstroke muscles in the domestic pigeon (Columba livia) during flight. , J. Exp. Biol 134, 1-16.
- 25.Dial K P, Biewener A A. (1993) Pectoralis muscle force and power output during different modes of flight in pigeons (Columba livia). , J. Exp. Biol 176, 31-54.
- 26.Pennycuick C J, Fuller M R, Oar J O. (1994) Falcon versus grouse: flight adaptations of a predator and its prey'. , Journal of Avian Biology 25, 39-49.
- 27.Poore S O, Ashcroft A, Sanchez-Haiman A. (1997) The contractile properties of the M. supracoracoideus in the pigeon and starling: A case for long-axis rotation of the humerus. , J. Exp. Biol 200, 2987-3002.
- 28.Bishop C M. (2005) Circulatory variables and the flight performance of birds. , J. Exp. Biol 208, 1695-1708.
- 30.Marshall R. (1990) Management of pigeon diseases. Proceedings Association of Avian Veterinarians 122-135.
- 31.Vereecken M, P de Herdt, Ducatelle R. (1998) Adenovirus infections in pigeons: A review. Avian Pathology. 27(4), 333-338.
- 32.Paulik S, Paulikova I, Levkutova M. (2004) Paramyxovirosis and sporting performance in racing pigeons. Slovensky Veterinarsky Casopis. Vydava Vyskumny ustav Veterinarnej Mediciny, Kosice. , Slovakia 29(6), 28-29.
- 33.Doneley B. (2008) Pigeons: Respiratory disease. In:. , Gloucester, UK:, Chitty J and Lierz M (eds). Manual of Raptors, Pigeons and Passerine Birds. BSAVA 320-327.
- 34.Pees M. (2008) Pigeons: Gastrointestinal tract disease. In Chitty J and Lierz M (eds). Manual of Raptors, Pigeons and Passerine Birds. BSAVA , Gloucester, UK: 328-333.
- 35.Pennycott T. (2008) Pigeons: Infectious diseases. In Chitty J and Lierz M (eds). Manual of Raptors, Pigeons and Passerine Birds. BSAVA , Gloucester, UK: 311-319.
- 36.Atkinson G, Nevill A M. (2001) Selected issues in the design and analysis of sport performance research. , Journal of Sports Sciences 19(10), 811-827.
- 38.In Chitty J, Lierz M.. Manual of Raptors, Pigeons and Passerine Birds. BSAVA , Gloucester, UK: 299-304.
- 39.Rothe H J, Nachtigall W. (1987) Pigeon flight in a wind tunnel. II. Gas exchange and power requirements. , J. Comp. Physiol 157, 99-109.
- 40.Pennycuick C J, Klaassen M, Kvist A. (1996) Wingbeat frequency and the body drag anomaly: Wind-tunnel observations on a thrush nightingale (Luscinia luscinia) and a teal (Anas crecca). , J. Exp. Biol 199, 2757-2765.
- 41.Pennycuick C J. (1997) Actual and “Optimum” flight speeds: Field data reassessed. , J. Exp. Biol 200, 2355-2361.
- 43.U.S. Department of Transportation, Federal Aviation Administration Flight Standards ServicePilot’s Handbook of Aeronautical Knowledge. FAA-H-8083-25A
- 45.Ohlson D. (2010) The Mathematics of Aircraft Navigation. Resource by Royal Academy of Engineers. www.raeng.org.uk
- 46.Bowdler D, Bullmore A J, Davis R A. (2009) Prediction and assessment of wind turbine noise – agreement about relevant factors for noise assessment from wind energy projects. , Institute of Acoustics, Acoustics Bulletin 34(2).
- 48.Pennycuick C J. (1982) The Flight of Petrels and Albatrosses (Procellariiformes), Observed in South Georgia and its Vicinity. , Philosophical Transactions of the Royal Society B: Biological Sciences 300(1098), 75-106.
- 50.Scullion F T, Scullion M G. (2007) Pathologic findings in racing pigeons (Columba livia domestica) with ‘Young bird sickness’. , Journal of Avian Medicine and Surgery 21(1), 1-7.
- 51.Hewinson R G, Griffiths P C, Bevan B J. (1997) Detection of Chlamydia psittaci DNA in avian clinical samples by polymerase chain reaction. , Vet. Microbiol 54, 155-166.
- 52.Scullion F T, Scullion M G. (2003) Investigation of diseases in young racing pigeons. In Scullion FT and Bailey TA (eds). Proceedings of World Association of Wildlife Veterinarians, Wildlife Session 27th World Veterinary Congress , Tunisia. WAWV. Ballygawley, UK: 73-75.
- 53.Scullion F T, Scullion M G. (2010) Multiresistant Escherichia coli in racing pigeons. The Veterinary Record. 167(22), 880.
- 54.P De Herdt, Ducatelle R, Lepoudre C. (1995) An epidemic of fatal hepatic necrosis of viral origin in racing pigeons (Columba livia). , Avian Pathol 24, 475-483.
- 55.Weissenböck H, Fuchs A. (1995) Histological and ultrastructural characterisation of hepatic intranuclear inclusion bodies in psittacine birds and pigeons. , Avian Pathology 24, 507-521.
- 56.Scullion F T, Scullion M G. (2010) Racing Pigeons In Meridith A and Johnston Delaney C (eds). Manual of Exotic Pets.
- 58.Miller W. (2008) Statistics and measurement using OpenStat. http://www.statpages.org/miller/openstat/
- 59.Henderson I, Parrott D, Moore N. (2004) Racing pigeons – impact of raptor predation. Report to Scottish Natural Heritage and Scottish Homing Union. Central Science Laboratory.
- 60.Pratt J G. (1955) An investigation of homing ability in pigeons without previous homing experience. , J. Exp. Biol 32, 70-83.
- 61.Kramer G, St Paul UV. (1952) . Heimkehrleistung-en von Brieftauben ohne Richtungsdressur. Verh. dtsch. zool. Ges , Wilhelmshaven .
- 62.Graue L C. (1965) Experience effect on initial orientation in pigeon homing. , Animal Behaviour 13(1), 149-153.
- 63.Dell'Ariccia G, Constantini D, Dell'Omo G. (2009) Waiting time before release increases the motivation to home in homing pigeons (Columba livia). , J. Exp. Biol 212(20), 3361-3364.
- 64.Michener M, Walcott S. (1967) Homing of single pigeons – analysis of tracks. , J. Exp. Biol 47(1), 99-131.
- 65.Perry A K, Blickhan R, Biewener A. (1988) Preferred speeds in terrestrial vertebrates: Are they equivalent?. , J. Exp. Biol 137, 207-219.
- 66.Norberg U M. (1990) Vertebrate Flight: Mechanics, physiology, morphology, ecology and evolution. , Berlin, Zoophysiology Series 27.
- 67.Norberg U M. (1996) Chapter 7. Energetics of flight. In: Avian Energetics and Nutritional Ecology. Carey C (ed.) , New York, US 199-249.
- 68.Wolf M. Lunds University (2009) Catching the Invisible: Aerodynamic Tracks and Kinematics of Bat and Bird Flight. PhD Thesis
- 69.Schnell G D, Hellack J J. (1979) Bird flight speeds in nature: Optimized or compromise?. , The American Naturalist 113(1), 53-66.
- 70.Kvist A, Lindström Å, Green M. (2001) Carrying large fuel loads during sustained bird flight is cheaper than expected. , Nature 413, 730-732.
- 71.Pennycuick C J, Battley P F. (2003) Burning the engine : A time-marching computation of fat and protein consumption in a 5420-km non-stop flight by great knots, Calidris tenuirostris. , Oikos 2, 323-332.
- 72.Tieleman B I, Dijkstra T H, Klasing K C. (2008) Effects of experimentally increased costs of activity during reproduction on parental investment and self-maintenance in tropical house wrens. , Behavioral Ecology 19, 949-959.
- 73.Xirouchakis S M, Andreou G. (2009) Foraging behaviour and flight characteristics of Eurasian griffons Gyps fulvus in the island of Crete. , Greece. Wildlife Biology 15, 37-52.
- 74.Biro D, Meade J, Guilford T. (2004) Familiar route loyalty implies visual pilotage in the homing pigeon. Proceedings of the National Academy of Sciences of the United States of America 101(50), 17440-17443.
- 75.Meade J, Biro D, Guilford T. (2005) Homing pigeons develop local route stereotypy. Proceedings Biological Sciences. The Royal Society 272(1558), 17-23.
- 76.Schiffner I, Fuhrmann P, Wiltschko R. (2013) Homing flights of pigeons in the Frankfurt region: The effect of distance and local experience. , Animal Behaviour 86(2), 291-307.
Cited by (4)
This article has been cited by 4 scholarly works according to:
Citing Articles:
Frontiers in Veterinary Science (2025) OpenAlex Semantic Scholar Crossref
Animals (2023) OpenAlex Semantic Scholar Crossref
Journal of Veterinary Healthcare (2018) OpenAlex