Concomitant in Vivo Voltammetric and Electrophysiological Analysis Indicate that Nociceptin/Orphanin FQ Affects Dopamine and then Serotonin Activities in Brain Substancia Nigra.
Abstract
Nociceptin/orphanin-FQ (NOCI) together with its receptor NOP are widely expressed in cortical and subcortical motor areas and it is known that NOCI acts as an anxiolytic attenuating the behavioral inhibition of animals acutely exposed to stressful/anxiogenic conditions.
Influence of NOCI upon the dopaminergic system has been observed in the ventral tegmental area and in the nucleus accumbens as well as an inhibitory action of NOCI is described upon serotoninergic mechanisms at cells and terminal levels. In particular, it is known that serotoninergic fibers from the raphe system project to the substancia nigra (SN) and that this modulation is behaviourally relevant.
In the present work, the effect of exogenous NOCI injected into the SN upon DA and 5-HT levels have been analyzed by means of differential pulse voltammetry and nafion-carbon fiber microelectrodes. Electrophysiological monitoring of multicell activity was concomitantly performed with the same microsensor.
It appeared that both levels of these biogenic amines were specularly altered, with possibly a driving influence of the DA activity upon the serotoninergic function(s).
Article Information
- Received
- Accepted
- Published
Academic Editor: Mozhgan Torabi, PhD. in Animal Physiology, Department of Biology, Faculty of Science, Shahid Chamran University of Ahvaz, Iran.
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2019 Francesco Crespi
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Corresponding author: Francesco Crespi, Biology, GSK, Verona, Italy —
Competing Interests
The authors have declared that no competing interests exist.
Funding
No specific funding statement was provided by the authors.
Data Availability
No data-availability statement was provided by the authors.
Acknowledgements
To M. Manzalini for skillful technical assistance and to Prof. P. Nuthall for english language
Citation:
Introduction
Nociceptin/orphanin-FQ (NOCI) is an opioid-like neuropeptide that activates a G-protein coupled receptor: the NOP receptor 1 NOCI and its receptor are widely expressed in cortical and subcortical motor areas 2.
In 1997 Jenck and Coll. 3 demonstrated that NOCI acts as an anxiolytic, attenuating the behavioral inhibition of animals acutely exposed to stressful/anxiogenic conditions although the anxiolytic mechanism of NOCI is at present not completely clarified.
It has been reported that treatment with NOCI reduces the firing activity of dopamine (DA) cells in the ventral tegmental area (VTA) 4and inhibits DA release in the nucleus accumbens 5. This is resulting in altered regulation of motor control 1, 6.
Concerning serotonin (5-HT), it is known that serotonergic fibers from the raphe system project to the substancia nigra (SN) and that this modulation is behaviourally relevant 7. In particular, an inhibitory action of NOCI is described upon serotoninergic mechanisms exerted at two different levels:
1. On dorsal raphe nucleus (RDN) neurons, where NOCI causes inhibition by increasing K+ conductance 8, and
2. On cortical serotoninergic nerve terminals, where NOCI inhibits 5-HT release 9.
In the present work, the influence of NOCI upon DA and 5-HT release in SN is analyzed in vivo, in situ and in real time by means of electrochemical (voltammetric) experiments using nafion-coated carbon fiber microelectrodes (mCFE) for selective measurement of these two neurotransmitters 10. In addition, the same mCFE is used for concomitant electrophysiology and voltammetric measurements in SN as described earlier 11.
Methods and Results
Animals
Experiments were performed on adult male Wistar rats (260-300 g, Harlan, Italy). All procedures were carried out in accordance with the Italian law (Legislative Decree no.116, 27 January 1992), which acknowledges the European Directive 86/609/EEC, and were fully compliant with GSK policy on the care and use of laboratory animal and codes of practice.
In vitro experiments using differential pulse voltammetry (DPV) with mCFE have been performed to analyze at first the electrochemical characteristics of NOCI. The mCFE was prepared and DPV was performed as already described 10 (Figure 1).
Figure 1. Top left: schematic representation of the mCFE: the protruding active tip of the carbon fiber (diam. 12µm) can be cut under the microscope at a length of 200 250µm for DPV - electrophysiology measurements. Top right: the tip of the mCFE before (left) and after nafion coating as described earlier (Crespi et al., 1988). 
Download figure
In vitro data showed that this peptide (17amino acids) is not electroactive up to a concentration 1 µM dissolved in artificial cerebral spin fluid (aCSF) at pH 7.4. This is useful information as it allows performing local injection of the peptide in SN without alteration of the voltammetric analysis.
In vivo experiments have been performed in anesthetized (urethane 1.5g/kg i.p.) adult male rats with the nafion-mCFE stereotaxically inserted in the SN following Paxinos and Watson coordinates 12. A stainless-steel injector was placed into the SN near the mCFE (less than 1mm).
In control experiments two groups of rats (n=4 each) were treated with artificial cerebral spin fluid (aCSF: vehicle, 0.5µl), that was injected in SN in 8-10s. The values of DA and 5-HT remained constant during 90min. (Figure 2 and Figure 3).
Figure 2. TOP: effect of local injection of vehicle (aCSF 0.5µl), or NOCI 0.1nm, 1nm or 10nm, respectively on DA levels and BOTTOM: effect of local injection of vehicle (aCSF 0.5µl), or NOCI 0.1nm, 1nm or 10nm, respectively on 5-HT levels monitored in the SN with DPV and nafion mCFE. N=4 each treatment. Data are presented as % of basal pre-treatment levels ± sem. Stats: 2wANOVA, *p<0.05 versus control (vehicle), Dunnett test. 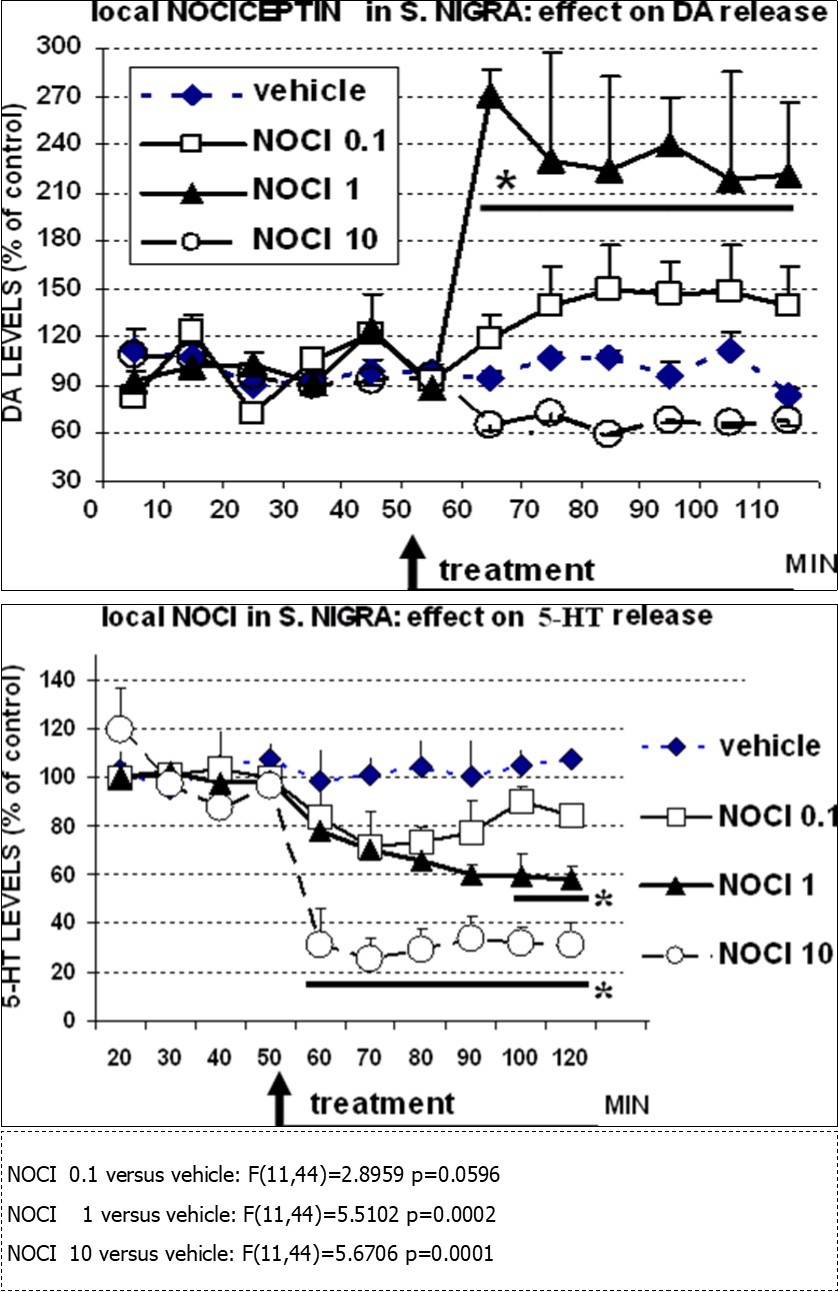
Download figure
Figure 3. Electrophysiological monitoring performed in the SN with the same nafion mCFE used for parallel DPV measurements as described earlier (Crespi 2002). Briefly, DPV scans last 30sec. and are repeated every 5min. Electrophysiology is then performed in the time gap between each DPV scan. In such a way, these two types of in vivo recordings could be performed concurrently in situ, allowing direct comparison of cell firing and monoamine release in the same brain region and in real time. In addition, the influence of pharmacological treatments upon both electrochemical and electrophysiological signals is therefore studied concurrently in real time and in the same animal. 
Download figure
In NOCI experiments local injection of NOCI (0.5µl, 0.1, 1 or 10 nanomoles, n=4 each dose) resulted in
A rapid, significant increase of DA levels in the SN up to 270% of control values within 10min then stabilizing at approximately 230% of control values. In particular, this was observed following local injection of NOCI 1 nanomole (nm), a significant increase to approximately 150% of control values was also obtained following local injection of NOCI 0.1 nm. In contrast, the highest dose (10 nm) was resulting in a decrease of the DA signal (Figure 2 top).
A dose-dependent, rapid decrease of 5-HT levels in the SN.
In particular, NOCI 10 nm was significantly decreasing 5-HT levels to approx. 30% of control values within 10min and NOCI 1nm to approx. 60% of control values within 40-50 min.
Parallel electrophysiological monitoring performed in the same animals with the same mCFE as described earlier 11 also indicated a dose-dependent, rapid decrease of (multi)-cell firing in the SN (Figure 3).
Discussion
The present original in vivo data demonstrates that NOCI locally injected in the SN directly affects cell firing as well as DA and 5-HT levels in this brain region. Taken together with the observations from Calo' et al 13 that NOCI is involved in:
(i) Inhibition of glutamate release/anti-epileptic action and disruption of spatial memory; (ii) Inhibition of serotonin release/anxiolytic action; (iii) Inhibition of mesolimbic dopaminergic transmission/anti-rewarding properties; (iv) Modulation of striatal dopamine and glutamate/effects on locomotor activity, the present data further support the wide role of NOCI in the CNS functions as a potent modulator of neurotransmitter activities. In particular, these data support the direct interaction of NOCI with the dopaminergic and the serotoninergic activities in the SN.
Indeed, for what concern dopamine, two parallel ex vivo approaches, dual in situ hybridization (ISH) and neurotoxic lesions of DA neurons by using 6-hydroxydopamine (6-OHDA) were applied in the ventral tegmental area (VTA) and substantia nigra pars compacta (SNc) by Norton et al. 14. This was done in order to verify whether NOCI acts directly on DA neurons i.e. upon NOCI-receptors (NOP), or indirectly by means of local circuitry, or both. It appears that NOCI mRNA was found largely on non-dopaminergic (i.e., GABA) neurons, whereas NOP mRNA was located on DA neurons. These data indicated that NOCI is in a position to influence DA neuronal activity by means of the NOP located on DA neurons. Accordingly, the present in vivo electrophysiological data support previous in vitro electrophysiological recordings observing that NOCI hyperpolarized the dopaminergic cells of the substantia nigra pars compacta and inhibited their firing activity 6.
Furthermore, in vivo microdialysis studies have shown a large increase of dopamine release (in the order of 300% of control values) in striatum when treating conscious rats with nociceptin at the micromolar concentration 15. This evidence is in accord with the present observation of a large increase of DA levels following local injection of NOCI in SN.
In contrast, intra-cerebroventricular administration of NOCI at a probe concentration of 1 mM but not at 0.1 mM significantly reduced rat nucleus accumbens dialysate DA levels in studies using a dual-probe microdialysis experimental design. Similar data were obtained when NOCI was applied to the ventral tegmental area of anesthetized rats by reverse dialysis while extracellular DA was sampled with a second dialysis probe in the nucleus accumbens 5. The observation that only the 1mM NOCI concentration but not the 0.1 mM concentration is acting upon DA levels may indicate that the technical approach used by these authors is not the most responsive. Nevertheless, this data is in accord with the present detection of reduced levels of DA in SN when the high dose of NOCI is injected (see Figure 2).
It is known that serotoninergic fibers from the raphe system project to the Substancia Nigra (SN) 16 and in particular that serotonin released from terminals in SN, derived from cell bodies in the raphe dorsalis nucleus, decreases the activity of the nigro-striatal dopaminergic system 17. It has been also observed that this modulation is behaviourally relevant 7.
Additionally, the present data showing large decrease of 5-HT levels in SN following local treatment with NOCI and a parallel dose-dependent, rapid decrease of (multi)-cell firing in the SN are in accord with results showing reduction of electrical activity of 5-HT neurons in RDN 18 as well as 5-HT release i.e. in cerebral cortex 19 after NOCI injection. Also, the inhibitory effect of NOCI on the nigral release of 5-HT can be related to the altered motor activity monitored i.e. using the fixed-speed rotarod test 6.
When considering the present data in a whole, it appears that there is a rapid effect of NOCI upon both DA and 5-HT signals in SN. The timing is similar although the effect is the opposite, with a very large increase of DA levels and a parallel large decrease of 5-HT values. However, while DA is rapidly largely significantly affected by NOCI 1nm, 5-HT is modified similarly by NOCI 10nm while is significantly less sensitive to NOCI 1nm (see Figure 2). Based on this observation one may propose that the rapid, large effect of NOCI 1nm on DA release may be possibly the driver of the comparatively slower and reduced change of serotonin levels, indicating that is dopamine that may be primarily influenced by NOCI in the SN.
The interaction between these two aminergic systems has been already reported in such diverse functions as temperature regulation 20, sleep 21, sexual behavior 22 and extrapyramidal function 23 although many results are conflicting 24.
In particular, it has been already reported that electrical stimulation of SN resulted in a rapid increase of the catecholaminergic DPVoltammetric signal followed by a slower decrease of the serotoninergic peak, both recorded simultaneously in the rat striatum 25. Accordingly, Kuhr and coll. 26 observed a rapid rise of DA levels in the caudate after electrical stimulation in SN. A similar and more consistent effect was observed when stimulating the medial forebrain bundle (MFB) and recording DA and 5-HT activities in the striatum 25, 27.
In conclusion, this work provides further evidence that DPV combined with mCFE is a valuable tool in the study of the in vivo effect of NOCI, an endogenous neuropeptide involved in a number of biological actions 28, 29, 30. In particular, it proposes a multifaceted implication of this neuropeptide on both dopaminergic and serotoninergic functions in the SN, suggesting a primary influence upon dopamine activity followed by the serotoninergic response. This data may be of importance in the interpretation of the biological functions of NOCI as indeed the NOCI-NOP receptor system is widely represented throughout the CNS 31.
For instance, the influence upon motor behavior may be considered as an implication of both DA and 5-HT in physiological as well as pathological conditions i.e. Parkinson disease (PD) 32.
Furthermore, the involvement of serotonergic mechanisms in the development of Levodopa-induced dyskinesias (LIDs) via aberrant processing of exogenous levodopa and release of dopamine as a false neurotransmitter in PD patients has been recently confirmed 33.
Additionally, the implication of NOCI within the regulation of feeding, body weight homeostasis, stress, the stress-related psychiatric disorders of depression and anxiety, and in drug and alcohol dependence, pathological situations involving both DA and 5-HT, has been described (for a review see 34).
Therefore, the present data proposing a complex interaction between NOCI, DA and 5-HT systems may be of help in the interpretation of physiological as well as pathological states and consequent development of therapeutical approaches.
References
- 1.Cox B M, Chavkin C, Christie M J, Civelli O, Evans C.et al.(2000) Opioid Receptors. In: The IUPHAR Compendium of Receptor Characterization and Classification. (Girdlestone D ed):. , London
- 2.Darland T, Heinricher M M, Grandy D Y. (1998) Orphanin FQ/nociceptin: a role in pain and analgesia, but so much more. Trends Neurosci.,21. 215-221.
- 3.Jenck F, Moreau J L, Martin J R, Kilpatrick G J, Reinscheid R K. (1997) Orphanin FQ acts as an anxiolytic to attenuate behavioral responses to stress. , Proc. Natl. Acad. Sci. U.S.A 94, 14854-14858.
- 4.Zheng F, Grandy O K, Johnson S W. (2002) Actions of orphanin FQ/nociceptin on rat ventral tegmental area neurons in vitro. , Br. J. Pharmacol 136, 1065-1071.
- 5.Murphy N P, Maidment N T. (1999) Orphanin FQ/nociceptin modulation of mesolimbic dopamine transmission determined by microdialysis. , J. Neurochem 73, 179-186.
- 6.Marti M, Mela F, Veronesi C, Guerrini R, Salvadori S. (2004) Blockade of Nociceptin/Orphanin FQ Receptor Signaling in Rat Substantia Nigra Pars Reticulata Stimulates Nigrostriatal Dopaminergic Transmission and Motor Behavior. , Journal of Neuroscience,doi:https://doi.org/10.1523/JNEUROSCI.0987-04.2004 24(30), 6659-6666.
- 7.Dray A, Davies J, Oakley N R, Vellucci S TongroachP. (1978) The dorsal and medial raphe projections to the substantia nigra in the rat: electrophysiological, biochemical and behavioural observations. , Brain Research 151(3), 431-442.
- 8.Vaughan C W, Christie M J. (1996) Increase by the ORL1 receptor (opioid receptor-like1) ligand, nociceptin, of inwardly rectifying K conductance in dorsal raphe nucleus neurones. , Br. J. Pharmacol 117, 1609-1611.
- 9.Werthwein S, Bauer U, Nakazi M, Kathmann M, Schlicker E. (1999) Further characterization of the ORL1 receptor-mediated inhibition of noradrenaline release in the mouse brain in vitro. , Br. J. Pharmacol 127, 300-308.
- 10.Crespi F, Martin K F, Marsden.CA (1988a) Measurement of extracellular basal levels of serotonin in vivo using nafion-coated carbon fibre electrodes combined with differential pulse voltammetry. Neuroscience27 885-96.
- 11.Crespi F. (2002) In vivo voltammetry and concomitant electrophysiology at a single biosensor to analyse ischaemia, depression and drug dependence. , Journal of Neuroscience Methods 119, 173-184.
- 13.Calo' G, Guerrini R, Rizzi A, Salvadori S, Regoli D. (2000) Pharmacology of nociceptin and its receptor: a novel therapeutic target.British. , Journal of Pharmacology129,1261-1283
- 14.Norton C S, Neal C R, Kumar S, Akil H, Watson S J. (2002) Nociceptin/orphanin FQ and opioid receptor-like receptor mRNA expression in dopamine systems. https://doi.org/10.1002/cne.10154 , J. Comp. Neurol.444: 358-368.
- 15.Konya H, Masuda H, Itoh K, Nagai K, Kakishita E et al. (1998) Modification of dopamine release by nociceptin in conscious rat striatum. , Brain Research 788(12), 341-344.
- 16.Dray A, Gonie T J, Tanner T. (1976) Evidence for the existence of a raphe projection to the substancia nigra of the rat . , Brain Res 113, 45-57.
- 17.Fibiger H C, Miller J J. (1977) An anatomical and electrophysiological investigation on the 5-HT projection from the DRN to the SN in the rat. , Neurosci 2, 975-987.
- 18.Vaughan C W, Connor M, Jennings E A, Marineih S, Alien R G et al. (2001) Actions of nociceptin/orphanin FQ and other prepronociceptin products on rat rostral ventromedial medulla neurons in vitro. , J. Physiol.534 849-859.
- 19.Mela F, Marti M, Ulazzi L, Vaccai E.Zucchini S et al.(2004) Pharmacological profile of nociceptin/orphanin FQ receptors regulating 5-hydroxytryptamine release in the mouse neocortex. , Eur. J. Neurosci.19 1-8.
- 20.Feldberg W, Lotti V J. (1967) Temperature response to monoamines and an inhibitor of MAO injected into the cerebral ventricles of rats. , Br. J. Pharmacol. Chemother 31, 152-161.
- 22.Gessa G L, Tagliamonte A. (1974) Possible role of brain serotonin and dopamine in controlling male sexual behaviour. , Adv. Biochem. Psychopharmacol 11, 217-228.
- 23.Hornykiewicz O. (1974) Neurohumoral interactions and basal ganglia function and dysfunction. In M.D. , Yahr (Ed.), The Basal Ganglia,Raven Press,New York 55, 269-278.
- 24.Lloyd K G. (1978) Neurotransmitter interactions related to central dopamine neurons. In. Essays in Neurochemistry and Neuropharmacology , Wiley,New York 3, 129-208.
- 25.Crespi F, Martin K F, Marsden.CA (1988b) Simultaneous in vivo voltammetric measurement of striatal extracellular DOPAC and 5-HIAA levels: Effect of electrical stimulation of DA and 5HT neuronal pathways. , Neuroscience Letters 90, 285-291.
- 26.Kuhr W G, Ewing A G, Caudill W L, Wightman R M. (1984) Monitoring the Stimulated Release of Dopamine withIn VivoVoltammetry. I: Characterization of the Response Observed in the Caudate Nucleus of the Rat j. , Neurochem 43, 560-569.
- 27.Crespi F, Paret J, Keane P E, Morre M. (1984) An improved differential pulse voltammetry technique allows the simultaneous analysis of dopaminergic and serotonergic activities in vivo with a single carbon-fibre electrode. , Neurosci. Lett 52, 159-164.
- 28.Meunier J C, Mollereau C, Toll L, Suaudeau C, Moisand C. (1995) Isolation and structure of the endogenous agonist of opioid receptor-like ORLi receptor. Nature377 532-535.
- 29.Reinscheid R K, Nothacker H P, Bourson A, Ardati A, Henningsen R A. (1995) Orphanin FQ: a neuropeptide that activates an opioid-like G protein-coupled receptor. Science270 792-794.
- 30.Mogil J S, Pasternak G W. (2001) The molecular and behavioral pharmacology of the orphanin FQ/Nociceptin peptide and receptor family. Pharmacol. Rev.53 381-415.
- 31.Slowe S J, Clarke S, Lena I, Goody R J, Lattanzi R. (2001) Autoradiographic mapping of the opioid receptor-like 1 (ORL1) receptor in the brains of mu-, delta- or kappa-opioid receptor knockout mice. Neuroscience106 469-80.
- 32.Politis M, Oertel W H, Wu K, Quinn N P, Pogarell O. (2011) Graft‐induced dyskinesias in Parkinson's disease: High striatal serotonin/dopamine transporter ratio. , Movement Disorders 26(11), 1997-2003.
Cited by (2)
This article has been cited by 2 scholarly works according to:
Citing Articles:
Neuroscience and Neurological Surgery (2022) OpenAlex
Journal of new developments in Chemistry (2020) OpenAlex Crossref