Abstract
The rise of epigenetics provides a new idea for studying the regulation of chronic pain-related genes and synaptic plasticity. External environmental stimuli can regulate BDNF genes through different epigenetic modifications. The epigenetic changes of the BDNF gene can affect the expression of its mRNA and protein and participate in the development of chronic pain. By reviewing the literature, this paper reviews the mechanism of epigenetic regulation of brain-derived neurotrophic factor (BDNF) in chronic pain, which provides some new directions and targets for the treatment of chronic pain.
Author Contributions
Academic Editor: Farah Yousef, Damascus University, Syria.
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2019 Xiang Wei, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Competing interests
The authors have declared that no competing interests exist.
Citation:
Chronic pain
Pain is an unpleasant subjective feeling and emotional experience associated with tissue damage or potential damage1. It is the fifth largest clinical sign in clinical practice and one of the most common clinical symptoms. Pain caused by peripheral tissue injury or disease Inappropriate treatment or treatment options and medications are inappropriate, Acute pain often develops into chronic pain(lasting more than 6 months).Chronic pain is a clinical syndrome characterized by pain, which is caused by a combination of biological, psychological and social factors. People with chronic pain are more than 20% in developed countries 2,and there are hundreds of millions of pain patients in China alone. Epidemiological surveys show that the prevalence of chronic pain in the elderly is 70 to 85%, and that of middle-aged people is 25 to 40%. With the aging of the population in China, pathological pain syndrome is more common among the elderly population, More than half of the population over 65 years old in China suffer from pain. Chronic pain lasts for a long time, stubborn and difficult to cure, therefore some negative emotional factors gradually breed, among which anxiety and depression occur. Epidemiological data show that 34 to 66 % of depression patients are accompanied by chronic pain, and up to 52% of patients with chronic pain have major depression 3.Therefore, the search for effective treatment methods and drugs to improve the quality of life of patients and reduce the socio-economic burden is of great significance
The pathogenesis of chronic pain is complex. Previous research hotspots have focused on the mechanism of neuropathic pain from the perspective of molecular proteinology and signal transduction pathways. Now the genetic mechanism of neuropathic pain and the study of epigenetic mechanisms are receiving increasing attention. Numerous studies have shown that the production and maintenance of neuropathic pain is closely related to changes in the transcription and expression levels of pain-related molecules and receptor genes on primary sensory neurons, secondary sensory neurons, immune cells, and glial cells. How many pain-related genes are procedurally initiated, transcribed and translated, increased in expression, or specifically inhibited, decreased in expression, thereby regulating pain, has now become a new direction of pain mechanism research, which involves the appearance of Genetics. The epigenetic changes are reversible, and the use of corresponding epigenetic interventions can re-express silent analgesic genes or reduce the expression of up-regulated pain-generating genes, providing new insights and Direction into the treatment of neuropathic pain.
Epigenetic Regulation
A large body of research evidence indicates that pain-related molecular changes after neurological damage are regulated by epigenetics. These genetic mechanisms are reversible modifications in the absence of changes in the DNA sequence. the heritability of the expression of the gene changes4. Epigenetic regulation refers to the structural adjustment of pretranscription genes at the chromatin level. It is a unique regulatory mechanism of the eukaryotic genome. Therefore, epigenetic regulation is also known as chromatin based gene expression regulation 5.Epigenetics is the result of interaction between environmental factors and genetic material in cells. It is a dynamic bridge between environmental stimuli and genotypes. Environmental factors change the function of genes by acting on key enzymes. Epigenetic mechanisms have a lasting effect on genes that are activated in a neuralgia environment. Currently, DNA methylation, histone modification, and non-coding RNA regulation play a vital role in gene activation and inactivation as well as phenotypic metastasis6, 7.
The formation of chronic pain requires a large number of genes to participate in it. Recent studies have reported that the gene expression of 12.4 % in chronic neuralgia rats is increased and 7 % is decreased 8.The products of these genes can regulate the neural network adaptability required for chronic pain production in a long-term and stable manner, and continuously and stably change gene expression and heritability, which are the two most important features of epigenetic regulation. Thus, changes in gene transcription activity in epigenetic regulation mediate the production and maintenance of chronic pain. Therefore, the rise of epigenetics provides a new idea for the study of chronic pain-related gene regulation 9. At present, epigenetic modifications that have received attention in the field of pain and analgesia mainly include: DNA methylation, non-coding RNA regulation, and histone modification, etc. They regulate pain-related molecular, Synaptic, and nuclear network activities in nervous system, producing different painful behavior phenotypes, including neuralgia, somatic pain, and visceral pain. (Figure 1)
Figure 1. Overview of epigenetic modification (SM Geranton, Current opinion in pharmacology, 2012)
Brain Derived Neurotrophic Factor
Brain derived neurotrophic factor (BDNF) is another vital neurotrophic factor discovered after nerve growth factor (NGF). It can maintain the normal physiological functions in the central and peripheral nervous system, plays an important role in the survival, differentiation, development of neurons, and can promote the regeneration of injured neurons10. With the deepening of research, BDNF, as a signal molecule connecting neuronal-glial intercellular information transmission, has attracted more and more attention in the development of pain11. In recent years ,A large number of studies have found that epigenetic modifications play an important role in the expression of BDNF gene. In the central system, the BDNF gene is the first recognized downstream target gene under epigenetic regulation. These studies provide a new direction for the pathogenesis of chronic pain and provide a new target for drug therapy. This article reviews the study on epigenetic regulation of BDNF gene and chronic pain.
BDNF Gene and Chronic Pain
Structure of BDNF Gene
BDNF has a complex structure, which has been studied in depth12, 13. Many studies have shown that rodents and humans have substantial similarities. The rat BDNF gene contains 9 exons, Which consists of 8 non-coding exons and 1 exon encoding the protein. Any of the 8 non-coding exons can be selectively spliced with the coding exon and transcribed to form BDNF mRNA . The human BDNF gene is located on chromosome 11 and encodes 247 amino acids. The protein contains 11 exons, each of which can be independently cleaved into a coding sequence, in which 9 promoters are present on exons I, VII, VIII. And IX, all of which initiate the transcription process.The coding sequence forms a different BDNF transcript14 The study found that external environmental stimuli can regulate different promoters of BDNF through different epigenetic modifications, which can cause changes in mRNA expression of different exons and ultimately lead to changes in BDNF gene expression.
Human and rodent BDNF genes are contains nine exons (Figure 2), Which has their own promoter, producing a variety of different transcripts. Interestingly, all of their transcripts were translated into the same BDNF protein12, 13, 15, 16. Some exons of BDNF between humans and rodents are highly conserved 17. Each exon is regulated by its own unique promoter for the temporal and spatial specificity of BDNF expression in an activity-dependent manner.
Figure 2. Structure of human and rodent BDNF genes. The exon is a box and the intron is a line. The sequence number of the exon is represented as a Roman numeral, and the molecular weights of the exons and introns are represented as Arabic numerals. Two polyadenylation sites (poly A) are encoded exons. The start codon ATG of the marker transcription initiation is indicated by a red box. The exon IX region encoding the pro-BDNF protein is shown in green boxes.. There are also some exons, such as exons II and IX, with alternative splice donor sites (A, B, C, D) containing different transcript variants.18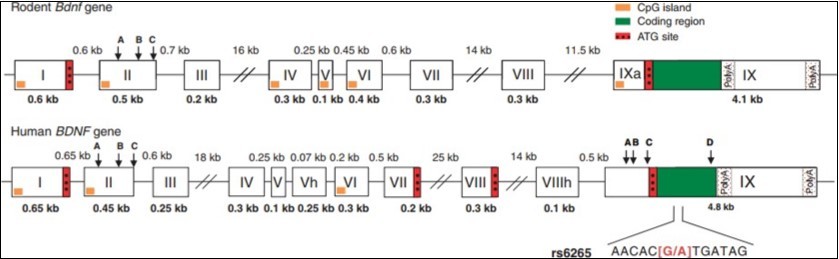
BDNF gene is Involved in the Development of Chronic Pain
Multiple patient and animal studies of chronic pain have shown that abnormal BDNF gene expression may be involved in the pathogenesis of chronic pain. Numerous studies have shown that BDNF is an important substance regulating nociceptive information, and participating in the occurrence and maintenance of pain sensitization 19. As early as 1999, the researchers found that increasing the concentration of BDNF in the brain increased the sensitivity of rats to pain, and the use of BDNF inhibitors significantly reduced the pain performance in rats20.In the CPIP model induced by rodent hind paw ischemia-reperfusion (IR) injury, microglia accumulate in the ipsilateral dorsal horn after ischemia for 3h, and the accumulated microglia release BDNF, increasing the excitability of neurons in the dorsal horn. And produce painful behavior 21.Studies have also found that spinal nerve ligation can increase the activity and number of neurons producing BDNF, and then the concentration of BDNF increases. This result indicates that the damage will increase the concentration and activity of endogenous BDNF 22. Other studies have also demonstrated this conclusion, such as temperature - induced hypersensitivity associated with elevated BDNF levels 23. The intensity of pain in chronic pancreatitis is directly proportional to the concentration of BDNF 24. These results indicate that pain does increase the expression of BDNF. During nociceptive stimulation in chronic pain, nociceptors are activated and release BDNF 25.The study found that the expression of BDNF was significantly up-regulated in the posterior horn of spinal cord in the pain-sensing (somatic pain) caused by cutting and the pain model (neural pain) caused by lumbar 5 root excision 26.In an animal model of neuropathic pain induced by nerve injury, increased expression of BDNF can be observed in intermediate cell neurons, large cell neurons, and their central terminals and spinal cord of dorsal root ganglia 27 intrathecal administration. BDNF scavenger TrkB/Fc can neutralize endogenous BDNF, thereby blocking its binding to receptors, and alleviating hyperalgesia induced by nerve injury 28. Geng et al found that early spinal cord dorsal rat spinal nerve ligation model The concentration of BDNF is significantly increased, combined with changes in the pain threshold of the rat, they believe that BDNF in the spinal dorsal horn is associated with the onset of neuropathic pain 29.Coull et al found that injection of BDNF into the spinal cord of normal mice induced symptoms of neuropathic pain in mice, whereas inhibition of BDNF mRNA expression or inhibition of BDNF and receptor by antisense RNA was first used. The combination and destruction of BDNF signaling process, peripheral nerve injury no longer induces neuropathic pain, this result clearly shows that BDNF takes part in the occurrence of neuropathic pain, its role is blocked will inhibit pain Occurs, and subsequent experiments further demonstrate the mechanism of action of BDNF in the pathogenesis of neuropathic pain 30.The above studies suggest that BDNF plays a key role in the generation and development of neuropathic pain.
BDNF is important for the regulation of pain, and epigenetics participates in the occurrence and development of pain. Recent research has focused on the epigenetic mechanism of the BDNF gene, which affects the BDNF gene by regulating environmental factors. Therefore, the lasting expression changes contribute to the production and development of neurological pain. The epigenetic changes of the BDNF gene can affect the expression of its mRNA and protein and participate in the development of chronic pain. However, there are few studies on how epigenetic modification regulates the expression of BDNF genes in pain. The following will be discussed further.
Regulation of Methylation on Expression of BDNF Gene and Chronic Pain
Histone and DNA Methylation
The methylation modification of BDNF gene has an important regulatory effect on the expression of this gene. Boulle et al.'s sequence analysis of the BDNF gene shows that human and rat BDNF gene promoter I, II, IV contain abundant CpG islands, The CpG island in the gene sequence is the main site of methylation modification. 31.A number of in vitro experiments observed changes in gene expression levels by changing the methylation status of the BDNF gene CpG island, It was found that the activity of the exon was reduced after methylation of several CpG sites on BDNF exon IV and transfection into mouse cortical neuron cells, However, the depolarization of cells with potassium chloride(KCl) can reduce the level of methylation of BDNF exons and increase the expression activity of the gene. 32Ishimaru et al. treated mouse neuronal cells with a DNA methyltransferase inhibitor 5-azo-2 indestruction- deoxycytosine, and compared with the control group found that the transcription of BDNF promoter I and IV increased33 These all experiments have shown that changes in the methylation state of the BDNF gene promoter or exon can affect the expression of the BDNF gene..
In-vivo experiments also showed that the methylation state of the gene is related to the expression of the BDNF gene. Studies have shown that repeated morphine injection may induce demethylation of two CpG sites in 197 and 176 of the BDNF exon IV gene promoter, which increases the expression of BDNF mRNA and protein levels in DRG neurons Moreover.
Intrathecalinjection of anti-BDNF antibodies can inhibit mechanical allergic reactions in rats after repeated morphine exposure. By increasing BDNF in DRG neurons, Inhibition of DNA methylation can significantly promote the production of opioid-induced hypersensitivity reactions. The results show that the methylation level of the BDNF gene exon in DRG neurons can regulate the expression of the BDNF, which is of great significance in opioid-induced hypersensitivity reactions. 34. Ikeqame and other studies found that BDNF methylation modification not only existed in brain tissue. It also exists in peripheral blood cells and saliva 35.Several studies have found changes in DNA methylation levels of the BDNF gene in the brain and peripheral blood of patients with chronic pain. A clinical study using blood to isolate genomic DNA found that women with myalgia syndrome showed significantly abnormal DNA methylation compared to age-appropriate health controls. Muscualgia syndrome is associated with different methylation of multiple genes, such as BDNF, HDAC4, etc.. 36.These studies have caused changes in the methylation level of animal BDNF gene by changing physical, chemical, and other environmental factors, thereby affecting the expression of BDNF gene The study found that there was a change in the level of total DNA methylation and related enzymes and binding proteins during the pain process. Expression changes of histone demethylase JMJD3, JMJD2A, transcription Suppressor MeCP2, histone dimethyltransferase G9a and DNA methylase DNMT in pain.
JMJD3
Recent studies have found that specific demethylation H3K27me2/3 produces deinhibitory histone demethylase JMJD3 also appears to be involved in the pain mechanism. It has been found that BDNF increases after peripheral nerve injury, leading to the occurrence of neuropathic pain. Neuropathic pain. Intrathecal administration of anti-BDNF antibodies inhibits thermal hyperalgesia and mechanical allodynia 37.JMJD3 can regulate the expression of BDNF in DRG neurons after nerve injury Generally,PRC2 silences the BDNF gene, and PRC2 contains the catalytic subbase EZH2. After stimulating neurons in vitro using NMDA, JMJD3 was recruited into the BDNF promoter to induce BDNF gene promoter demethylation .Therefore, the deinhibition of the promoter helps to increase the BDNF expression in mature neurons38.In addition to promoter inhibition, BDNF is also enhanced through the acetylation mechanism and through the role of CREB kinase / CBP39. In addition to its contribution to the pain process, JMJD3 also participates in the inflammation mechanism. JMJD3 increases after inflammation such as LPS.Studies have also been found that transcriptional regulation of Stat1 and Stat3 activates the expression of inflammation related genes in macrophages cultures of microglia. For example, Lee et al. described an increase in spinal cord injury(SCI) produced by JMJD3 in endothelial cells, which result in the elevation of cytokine IL-6 via demethylation of its promoter40.The incident has been confirmed in vitroand JMJD3 siRNA inhibits IL-6 expression in oxygen/glucose deprivation reactions. Therefore, the epigenetic modification of the BDNF promoter plays a vital role in the activation of neurons, and an increase in the level of BDNF may be observed after the injury of neurons .JMJD3 may be a Potential target for reducing neuropathic pain after neurotraumatic lesions by reducing inflammation and neurotrophic factor expression.
JMJD2A
JMJD2A, also known as KDM4A or JHDM3A, is a histone demethylase. It can catalyze the demethylation of H3K9 and H3K36 in the trimethylation / dimethylation state to different methylation levels(H3K9me2/3 and H3K36me2/3). In addition, JMJD2A can also act as a coactivator or cosuppressor binding protein of a transcription factor to regulate gene expression at the transcription level. The study found that the VPA treatment of rat embryonic neural stem cells led to a strong increase in the expression of BDNF mRNA, which was related to the demethylation of H3K9 at the Bdnf IV promoter in rats. It was further found that both KDM2A and KDM2C occupied the exons 5 of the rat promoter IV and BDNF genes under VPA conditions. At the same time, the reduction of the VPA-mediated H3K9me3 level was reduced by siKDM4A/C, while the level of H3K36me3 in the BDNF promoter did not change significantly. These results show that KDM2A/C is necessary for the increase of BDNF expression during neuron differentiation and is related to the demethylation of the promoter region H3K9me3. 41.In addition to participating in neuronal differentiation, spinal cord JMJD2A dependent histone demethylation also plays an important role in traumatic hypersensitivity reactions. Zhou et al. found that the mRNA and protein of JMJD2A were increased highly in neuronal neurological pain induced by CCI or SNI of the sciatic nerve. The JMJD2A participates in and maintains the development of neurotic pain by combining with the BDNF promoter and demethylated H3K9me3 and H3K36me3 on its promoter to upgrade the expression of BDNF. In addition, the knockout of JMJD2A blocked the hypersensitivity response of mouse with chronic pain caused by CCI and SNI. These results show that the increase of JMJD2A promotes the neurological pain of BDNF, which can treat as a hopeful target for the cure of chronic pain.42.
MECP2
The study found that there was a change in the level of total DNA methylation and related enzymes and binding proteins during the pain process. MeCP2, or methylated CpG binding protein, is the most clearly studied member of the currently methylated DNA binding protein(DNA-binding protein, MBD) family 43. In the nervous system, the BDNF gene is the first recognized downstream target gene regulated by MeCP2 44. In recent years, more and more evidence has shown that MeCP2 is closely related to the formation of chronic pain. MeCP2 binds to the CpG site of methylation on DNA to inhibit the transcription of specific genes, while MeCP2 after phosphorylation loses its binding ability. 45. The study found that the total level of DNA methylation and methylation of MeCP2 mRNA levels in the neurological pain model induced by chronic sciatic compression injury (CCI) were significantly increased. After the intervention of 5-azacytidine(5-azacytidine, 5-AZA), the total DNA methylation level and MeCP2 levels were reduced. At the same time, CCI induced mechanical hyperalgesia and thermal hyperalgesia were significantly relieved 46. MeCP2 is a protein related to Rett syndrome. Clinical studies have found that patients with Rett syndrome with MeCP2 mutation show insensitivity to pain. The binding pattern of MeCP2 on the genome also changed after nerve injury. The study found that MeCP2 expression in the central amygdala (CeA) increased and MeCP2 local silencing alleviated inflammatory pain in the state of sustained inflammatory pain, while the over-expressed MeCP2 mice showed pain sensitivity and a large number of gene expression changes such as BDNF in the central amygdala 47. Some studies have shown that the formation of polymers between MeCP2 and HDACs has an inhibitory effect on the expression of BDNF promoter IV. After depolarization of rat nerve cells in vitro experiments, it was found that the phosphorylation level of MeCP2 increased. However, increased phosphorylation of MeCP 2 can separate the binding sites of MeCP 2 and BDNF gene, thus relieving the inhibition of expression of BDNF gene. 48.In addition, DNA methylation in the prefrontal cortex and amygdala decreased after nerve damage, and this decrease is closely related to the degree of pain 49. This suggests that nerve injury induced DNA methylation in the central nervous system has mediated the development of pain.
In the nucleus accumbens, chronic stress increases the DNA differentiation of the neurotrophic factor(gdnf) gene promoter source and the binding of MeCP2. MeCP2 reduces the acetylation of the histone H3 on the gdnf promoter through interaction with HDAC2., This reduces gdnf gene expression and produces anxious behavior 50. Therefore, the relationship between DNA methylation and BDNF in chronic pain remains to be further explored.
Histone Dimethyl Transferase G9a
G9a(encoded by Ehmt2) is an H3K9 methyltransferase that belongs to a typical histone methyltransferase and is responsible for gene silencing. Recent studies have found that G9a is also closely related to pain formation. As mentioned above, methylated CpG binds methyl-CpG binding proteins, such as MeCP2. After MeCP2 binds to methylated DNA, it will raise transcription factor formation complexes such as G9a 51. Therefore, G9a may be an essential epigenetic marker protein for MeCP2 inhibition of gene transcription in chronic pain
In fact, a large number of pain-related genes, such as BDNF, are subject to the epigenetic regulation of G9a in many environments47.Zhang et al. found that the expression of G9a and H3K9me2 proteins in CeA central amygdala in mice with persistent inflammatory pain was reduced. The level of G9a on the BDNF gene promoter was also reduced and accompanied by an increase in BDNF protein levels. CeA local injection of G9a inhibitors reduced H3K9me2 and increased BDNF protein levels, and delayed the recovery time of persistent inflammatory pain 47. In addition, the pain threshold in mice with G9a transgenic, CeA local injection virus silent G9a expression, decreased.
A recent research showed that Mecp2 expression increased in the central amygdala (CeA) of mice due to CFA-induced chronic inflammatory pain and repeated morphine exposure. The increased MeCP2 binding and inhibition of transcription inhibition factor factor protein methyltransferase G9a resulted in a decrease in the inhibition marker H3K9me2 catalyzed by G9a in CeA, and inhibition of G9a increased the expression of BDNF.
DNMT
Recent studies have related DNA methylation of specific genes with pain allergic reactions. Current research has found that SNL increases the binding of DNMTs to BDNF promoter, which means SNL increases the spinal cord DNA methylation of BDNF promoter in rats. It is worth noting that although SNL increases the combination of DNMTs with BDNF promoters. At the same time, Tet1 dependent DNA demethylation of spinal cord was found to take part in the process of injury perception. Tetl mediates the conversion of 5-methylcytosine(5mC) to 5- hydroxymethylcytosine(5hmC), thereby promoting DNA demethylation. In the SNL rats model, the Tet1 expression was increased in the dorsal angular neurons and the CpG dinucleotide hydroxylation in the BDNF promoter was 5mC to 5hmC to promote the expression of the spinal cord BDNF after surgery. The concentration of Tet1 binding and 5hmC was reduced by focal knock of Tet1 expression in spinal cord. The concentration of 5mC in CpG site in BDNF promoter was further increased and the expression of BDNF in spinal cord was reduced, while the abnormal pain was reduced 52.
In addition, SNL upgrade Tet1 expression also inhibits the binding of DNA methyltransferase(DNMTs) to the BDNF promoter, which is a catalyst. The necessary conditions for the transcription silence of metron-5-cytosine(5C) to 5mC. In summary, these data show that at the CpG site of the BDNF promoter, SNL increased Tet1 expression promotes DNA demethylation by converting 5mC to 5hmC and inhibiting DNMT binding to regulate spinal cord BDNF expression, thus leading to the development of abnormal behavioral pain. (Figure 3 and Table 1).
Figure 3. When DNA methyltransferase (DNMTS) catalyzes the addition of cytosine at the 5' position to the methyl group, DNA methylation happened. When there is no DNA methylation, the transcription factor and RNA polymerase 2 (RNAP II) bind to DNA, leading to gene expression. Recent researches suggests that members of the TET family are likely to undergo active DNA demethylation. DNA methylation cause transcriptional repression by recruiting proteins with a methyl binding domain such as MeCP2, which can further recruit a co-inhibitor complex containing HDAC. TSS, transcription start site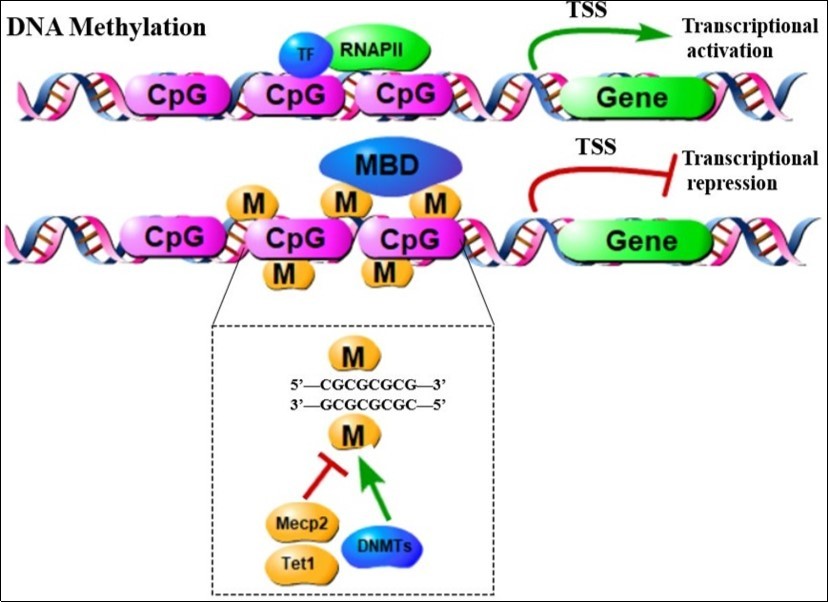
| Enzyme | Alteration | Molecular effect | Effect on geneexpression | Gene altered | Direct relation to pain | Reference |
| JIVLJD3 | increased | Demethylation of H3k27me2/3 | De-repression | BDNF,IL-6 | NO | Uchida,H et a1,2013; Ernest,P et a1,2016 |
| JMJD2A | increased | H3k9me3/H3k36me3 | Gene expression | BDNF,5-HT | NO | Zhou,J et a1,2017 |
| MECP2 | increased | Binding to CpG | Gene silencing | BDICF | YES | Wang,Y et a1,2011 Zhang,Z et a1,2014 |
| DNMT | increased | Binding to CpG | Gene expression | BDNF | YES | Hsieh, M et a1,2016 |
| Tetl | increased | Demethylation | Gene expression | BDNF, 5inc-51unc | YES | Hsieh, M et a1,2016 |
| G9a | decreased | H3k9ine2 | Gene silencing | BDNF | YES | Zhang, Z, et a1,2014 Ooi,L et a1,2007 |
Regulation of Histone Acetylation on Expression of BDNF Gene and Chronic Pain
Histone modification as an important epigenetic regulation also affects BDNF gene expression. Histone modifications include methylation, acetylation, phosphorylation and many other regulatory modes. Histone modifications can affect the binding of genes through the regulation of histone structure, leading to changes in gene expression. Some studies have shown that histone tails (methylation of H3 and H4), Modifications, acetylationwhich cause transcription of inflammatory molecules, such as cytokines and chemokines, are responsible for the development of chronic pain. In these cases, HAT appears to be involved in chemokine expression, and HDAC is associated with cytokine expression, which together lead to neuropathic pain.Next we mainly discuss the effect of histone acetylation on the BDNF gene.
In the chronic neuralgia model, acetylation occurs on many pain-related gene promoters of DRG, such as opioid receptors and bdnf genes 53. The increase of H3 and H4 acetylation on the BDNF gene promoter induced by nerve damage and inflammation, the expression of the BDNF gene was increased, and excitatory and inhibitory Synaptic transmission was involved, causing the production and maintenance of neuralgia. In the persistent inflammatory state, the expression of BDNF regulated by histone acetylation is increased, and chronic pain behavior is produced by activating TrkB and initiating Synaptic metastasis in the middle slit macronucleus(Glualyaji) via the PLC-PKC signal pathway54, 55. Neural injury promotes the increase of histone H3 and H4 acetylation in the BDNF promoter region in DRG and spinal cord injury to regulate the expression of BDNF, and can help induce or maintain nerve pain 56, 57
Ishimaru et al. treated mouse neuroblastoma cells with the HDACs inhibitor trichostatin, and discovered that the acetylation levels of histone H3 and H4 bound to BDNF promoter I were increased, while the expression of BDNF gene was also increased 58. Subsequent animal experiments have also dig out that elevated standard of histone acetylation are positively correlated with expression of BDNF 59, 60. Uchida H et al. showed that the initial increased expression of BDNF exon I controlled by epigenetic mechanisms may join in the development of neuropathic pain. In a partial sciatic nerve ligation model, they found that histone H3 and H4 acetylation regulate BDNF promoter transcription. Chromatin immunoprecipitation (ChIP) assay showed that genomic H3 and H4 acetylation of BDNF promoter I increased significantly on one day after nerve injury, and histone acetylation levels remained elevated for at least 7 days. Epigenetic control of protein acetylation is required for the induction and maintenance of BDNF promoter transcription in DRG 61. Similarly, studies have shown that mechanical hyperalgesia and elevated expression of BDNF/SYN1 in the spinal dorsal horn can be induced by inoculation of Walkers256 tumor cells in rat tibia. Intraperitoneal injection of the HDAC inhibitor, suberoylanilide hydroxamic acid (SAHA), reverses these changes and exhibits an antinociceptive effect. These findings suggest that regulation of histone acetylation levels is at least partially involved in BDNF-induced synaptic plasticity in an indirect manner, thereby inducing pain central sensitization in BCP rats.
Winston et al.62gave intermittent chronic stress stimulation to pregnant rats at a certain intensity, and found that female offspring presented strong and sustained intestinal hypersensitivity response to transient stress stimulation of this intensity as adults, which was stronger than male offspring. However, blocking BDNF protein receptor or knocking BDNF expression by small interfering RNA technology can reduce the intestinal hypersensitivity of female offspring, indicating that it is related to up-regulation of BDNF protein level in spinal dorsal horn. In addition, continuous administration of HAT inhibitor curcumin in female offspring can alleviate the stress related hypersensitivity of the newborn, while the mRNA and protein level of BDNF decreased, suggesting that HAT inhibitor may alleviate the intestinal hypersensitivity caused by chronic stress by inhibiting the expression of BDNF.
Curcumin has long been recognized as an inhibitor of HAT activity and used to treat of neuropathic pain. Studies have found that curcumin has an antinociceptive effect in a neuropathic pain model of chronic compressive injury (CCI) rats. The effects of curcumin on P300/CBP and BDNF was studied in a model of chronic compression injury. Treatment with curcumin for 7 consecutive days significantly attenuated CCI- induced thermal hyperalgesia and mechanical allodynia. Chromatin immunoprecipitation analysis showed that curcumin can reduce the recruitment of p300/CBP and acetyl histone H3/acetyl histone H4 to BDNF gene promoters. A similar result was also observed after curcumin treatment. These results indicate that curcumin have a therapeutic action in neuropathic pain by weakened BDNE gene expression mediated by p300/CBP HAT activity.63Another group studied the neurotic pain induced by HAT activity in rat CCI associated with transcription coactivator P300.They invented that the expression of P300 in the spinal cord was increased after CCI, and that intrathecal injection of the slow virus expressing P300 shRNA reduced the increase of P300 and reduced the increase of BDNF mRNA in the spinal cord. All the time, these reductions are followed with reductions in the hypersensitivity reactions caused by CCI. Intrathecal administration of the HAT inhibitor C646 reproduces the results. they further studied the p300/CBP, which stand for two functionally interchangeable proteins, expressed BDNF and COX2 in the spinal cord. Both the P300 and CBP are copositioned with BDNF or COX2 in the dorsal neurons of the spinal cord. CCI animals are known to have BDNF and COX2 increases in the spinal cord. Consistent with these facts, nerve damage causes P300, CBP, and H3K9ac, but not the increase in the binding of H4K5ac to the BDNF promoter, and all these proteins are combined with the BDNF promoter. Curcumin treatment prevented these expression, which is the same as the changes in the associated mRNA. Recent studies have also found that in the CCI model rats, the injection of HDAC inhibitor TSA into the middle slit large nucleus can increase the level of H3 acetylation on the NGF gene promoter, resulting in an increase in NGF protein expression, starting the NGF/Trk signaling pathway, which mediates the migration of U opioid receptors. Upper membrane Activated to produce highly effective analgesic effects.Does BDNF have the same mechanism? The relationship between the levels of histone modification in brain tissue or peripheral blood and the expression of BDNF in patients with chronic human pain has not been reported. (Figure 4 and Table 2).
Figure 4. In the chromosome, DNA is wrapped around a single histone to form the basic unit of DNA packaging - the nucleosome. Epigenetic modification on histones for histone translational histone acetyltransferase (HAT; acetylated histone), histone deacetylase (HDAC; removal of histone acetyl), histone methyltransfer Enzyme (HMT; methylated histone), histone demethylase (HDM; removal of histone methyl), enzymatic mechanism of protein kinase (PK; phosphorylated histone) and protein phosphatase (PP; removal) Histone phosphate group).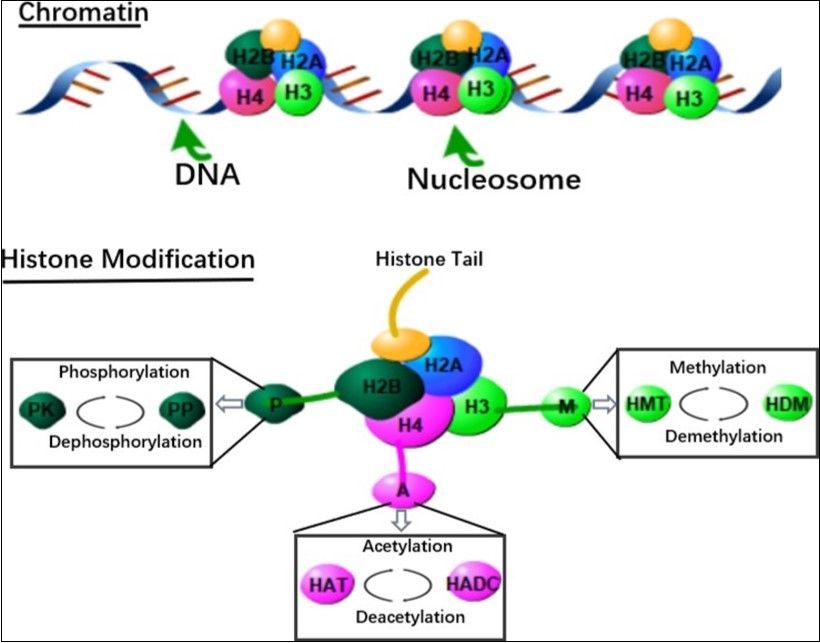
| Pain model | Enzyme | Inhibitor | Histone acetylation | Site | Gene altered | Reference |
| Visceral pain | HAT | Curcumin | SP | BDNF | Winston, et a1,2014 | |
| Neuralgia Pain | / | H31114 | DRG | BDNF | Uchida,et a1,2013; | |
| Inflammatory pain | / | / | H31114 | Central nucleus | BDNF | Uchida,et a1,2013; |
| SNL | / | / | H31114 | DRG | BDNF | Wang x, et al.2014 |
| BCIP | liDAC | SAHA | DRG | BDNF | Hoget a1,2016 | |
| CCI | HAT | Curcumin | H31114 | SP | P300/CBP/BDNF/ COX-2 | Zhu, X., et al.2014 |
| CCI | HAT | C646 | H3K9ac | SP | P300/CBP/BDNF/COX-2 | Zhu, I, et al.2014 |
| CCI | HDAC | TSA | 113 | Central nucleus | NGF | Tao, W., et a1.2016 |
MiRNA Regulation of BDNF Gene Expression and Chronic Pain
Neurological pain is chronic pain caused by somatosensory system diseases. MicroRNAs(miRNAs) have been extensively studied for the development of neurological pain and neuroinflammation caused by nerve damage. But, the specific mechanisms by which miRNAs are involved in neuropathic pain remains unknown.
The target miRNA of the BDNF gene can directly target the 3-UTR region of its target gene mRNA. The 3-end non-translation region(3 '-UTR) of the BDNF is an important binding region of the miRNA and recognizes the target mRNA through base complementary pairing., According to the different degree of complementarity, the silent complex degrades the mRNA of the target gene or deters the translation of the target gene, resulting in a change in the level of expression of the target gene. Multiple miRNAs have a regulatory effect on the BDNF gene. Studies have shown that the abnormal regulation of microRNA on BDNF is related to chronic pain. Genda et al. found that 111 types of miRNA expression were significantly regulated in CCI rats in the spinal cord dorsal after surgery 64.The study found that miRNA-195 inhibited the expression of BDNF gene 65. In the rat model of SNI, the expression of miRNA-195 in the DRG was increased 66, After the spinal nerve ligation, the expression of miRNA-195 in the spinal cord was increased and continued to be at least 14d 67.Up- regulation of miRNA-195 expression strengthened mechanical pain and pain allergies after peripheral nerve damage, and the opposite result was observed after the use of miRNA-195 inhibitors 67.It is speculated that miRNA-195 may participate in the development of chronic pain by regulating the expression of BDNF.
Studies have shown that miRNA-132 can indirectly affect the secretion of BDNF by regulating MeCP2 in the hippocampus of rats 68 Similarly, in the formalin induced acute pain rat model, miRNA-124a was lowered in the spinal cord, which was related to the increase of MeCP2 mRNA.69 Intravenous administration of miRNA-124a inhibitors aggravated pain behavior, and miRNA-124a mimics significantly attenuated formalin-induced inflammatory pain.69 In another study, intrathecal injection of miRNA-124 was used to treat persistent pain allergies induced by reversible corner fork gum and to prevent the occurrence of mechanical abnormal pain in the SNI model rats 70, Possible mechanism is to alleviate neuropathic pain by inhibiting the expression of BDNF mRNA and protein
More data on the effects of analgesic and pain-inducing miRNAs. miRNA-183 achieves analgesia by targeting the regulation of Nav1.3 and BDNF expression in DRG.
Intrathecal injection of lentivirus expressing miRNA-183 restored SNI-induced down- regulation of miRNA-183, resulting in decreased expression of Nav1.3 and BDNF mRNA in DRG and significantly attenuated SNL-induced mechanical allodynia in rats. 71 Neuropathic pain in rats CCI leads to down-regulation of miRNA-1 in the sciatic nerve. After CCI, miRNA-1-targeted pain-related proteins BDNF and Cx43 are up- regulated in the sciatic nerve and DRG, ipsilateral to the rat. Significant mechanical allodynia occurred in the hind paws 72.Lin et al found that the expression of miRNA- 183 in the dorsal root ganglia was significantly down-regulated after L5 spinal nerve ligation, and this was associated with the maintenance of mechanical allodynia. Furthermore, intrathecal injection of a lentivirus expressing miRNA-183 significantly attenuated mechanical allodynia and was associated with a significant downregulation of BDNF 73
In addition, CCI-induced rat hypersensitivity was reduced by the use of miRNA- 206 mimic, a synthetic double-stranded RNA mimicking endogenous miRNA-206, and miR-206 in rat dorsal root ganglia after CCI was down-regulated, while the expression of BDNF mRNA and protein was increased in vivo. miRNA-206 mimetic attenuates mechanical allodynia and thermal hyperalgesia by inhibiting the expression of BDNF mRNA and protein in CCI rats. These results indicate that miRNA-206 attenuates the development of neuropathic pain by targeting BDNF, indicating miRNA- 206 may be a potential target for the treatment of neuropathic pain 74 .
The study by Li Haiqin recorded for the first time the role of miRNA-375 / JAK2/STAT 3/BDNF signal conduction in the development of chronic morphine tolerance. Studies have shown that miRNA-375 can negatively regulate JAK2 to inhibit BDNF expression, thereby blocking the development of tolerance. The miRNA-375 level in the morphine tolerance model is reduced, and the JAK2 expression in chronic morphine therapy is negatively related. Lowering miRNA-375 levels can induce BDNF expression and induce behavior similar to pain allergies and spinal cord neuronal sensitivity, and ectopic expression of miRNA-375 or JAK2 participates in chronic morphine tolerance. The upward adjustment of miRNA-375 improved the morphine tolerance in the dorsal root ganglion of mice by inhibiting the JAK2/STAT 3 pathway, and the upward adjustment of miRNA-375 could inhibit the expression of BDNF by lowering the JAK2 in mice that tolerate morphine75. The study found that the miRNA-219 was reduced after the CFA injection in rats and the CpG island on the gene promoter was demethylated, and CaMKII γ was involved in CFA-induced pain by regulating the dorsal horn of the spinal cord. Studies have shown that miRNA-219 promotes the resistance to injury in mice DRG, followed by continuous morphine treatment. After chronic morphine treatment, the expression of miRNA-219 in the dorsal root ganglion of mice was reduced, and the expression of CaMKIl γ was increased. The expression of miRNA-219 and CaMKIl γ are closely related to the development of morphine tolerance. The expression of proteins and mRNA in BDNF in the DRG is induced by time-dependent methods through prolonged morphine exposure, which is subject to the transcriptional regulation of miR-219 and CaMKII γ. Removal of the DNF TrkB-Fccanpartially reducemorphine tolerance. Overexpression of miRNA-219 using miRNA-219 simulators or the reduction of CaMK1l γ expression using CaMK1l γ small interference RNA significantly delayed morphine tolerance. In addition, hyperalgesia and spinal cord neuron sensitization were caused by knockout miRNA-219 in control mice or functional inhibition of miRNA- 219, both of which were inhibited by CaMK1l γ small interference RNA or TrkB-Fc.76 These results prove that miRNA-219 promotes the development of chronic tolerance of morphine analgesia in the DRG of mice by targeting CaMKII γ and lift the expression of CaMKII γ dependent BDNF.
Alternatively, beneficial effects can be achieved by inhibiting pain-induced miRNAs. In the SNI model, the expression of miRNA-195 was up-regulated in DRG 77; the expression of miRNA-195 was up-regulated for at least 14 days after spinal nerve ligation. Up-regulation of miRNA-195 expression potentiated mechanical pain and hyperalgesia following peripheral nerve injury, whereas the opposite was observed with miRNA-195 inhibitors 78. The study found that miRNA-195 can regulate the expression of BDNF gene 79. It is speculated that miRNA-195 may participate in the development of chronic pain by regulating the expression of BDNF. In bone cancer induced pain model mice,it can induce an upward adjustment of p-CREB and CRTC1.
Adenovirus expression of CRTC1-siRNA by intrathecal injection effectively relieved mechanical pain and spontaneous pain caused by bone cancer by central inhibition of CRTC1, parallel to inhibition expressed by spinal cord BDNF and miRNA-212 / 132. Since the activation of the signal and the formation of a positive feedback loop of its target gene contribute to neuronal plasticity, an increase in CRTC1 can enhance the transcription of the CREB gene in the spinal cord and contribute to the maintenance of chronic pain.CRTCI-SiRNA weakens pain behavior induced by bone cancer and reduces the expression of CREB/CRTCI-target genes in the spinal cord, including BDNF and miRNA-212/132.80 Analgesia is also produced by inhibition of miRNA- 221, which is caused by up-regulation of cytokine signaling inhibitor 1 , a protein that inhibits cytokine signaling. After SNI, miRNA-221 is up-regulated in spinal microglia. Spinal cord miRNA-221 inhibitor down-regulates pro-inflammatory cytokines and reduces neuropathy-induced hypersensitivity 81 (Figure 5 and Table 3).
Figure 5. Epigenetic control of miRNA in pain state. A) Normal BDNF protein translaton. B) Functonof miRNA: mRNA degradaton and inhibiton of protein translaton. C) Changes in miRNA expression during pain: some increase, such as miR-195, and some decrease, such as miR-183, these changes cause pain by regulating the expression of BDNF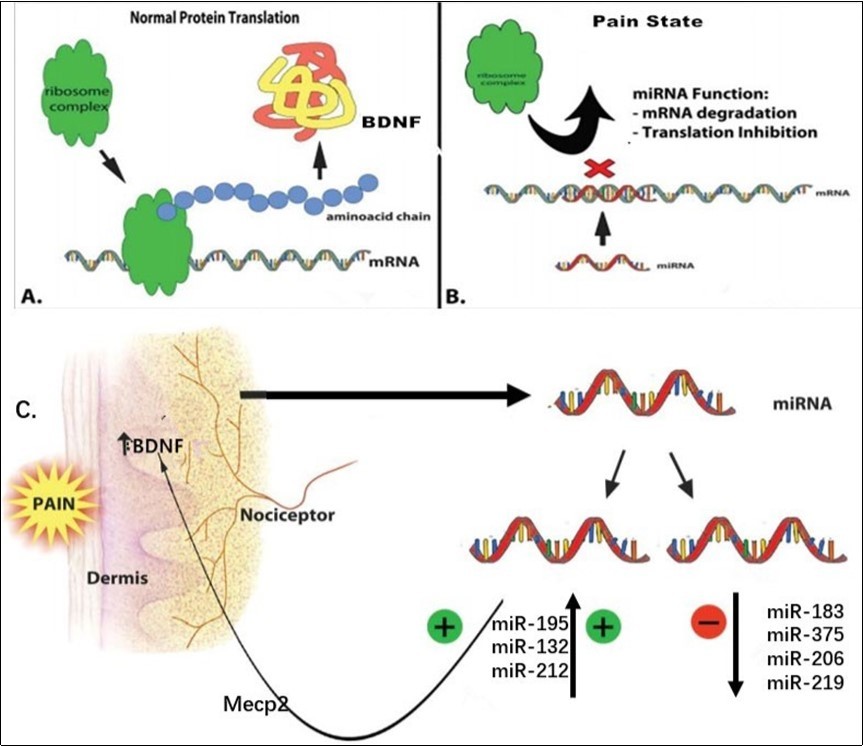
| miRNA | Alteration | Site | Gene altered | Reference |
| miR-206 | decreased | DRG | BDNF | Sun, et a1,2017 |
| miR-1 | decreased | Sciatic nerve | BDNF/Cx43 | Neumann,et a1,2015 |
| miR-183 | decreased | DRG | BDNF/Nav1.3 | Lin, et a1,2014 |
| miR-375 | decreased | DRG | BDNF/STAT3/JAK2 | Li,et a1,2017 |
| miR-219 | decreased | DRG | BDNF/CaMICIIT | Hu,et a1,2016 |
| miR-212/132 | increased | SP | BUNT CRTC' 'CREB NR2B | Ying,et a1,2016 |
| miR-195 | Increased | DRG | BDNF | Yu,et a1,2011 Shi,et a1,2013 |
Summary and Expectations
BDNF has a wide and important role in the function of the central nervous system. The study of the effect of epigenetic modification on the expression of BDNF genes has attracted more and more attention. As an important molecular control mechanism in organisms, epigenetic regulation closely links complex genetic factors with diverse environmental changes. In both animal and clinical studies, it was found that the regulation of BDNF gene by epigenetics was related to chronic pain.
Based on the literature of recent years, the epigenetic mechanism of BDNF gene regulation for chronic pain needs to be further studied. It mainly includes the following aspects:
(1) To clarify the effects of pain on the expression of BDNF genes in different brain regions, That is, under the pain state, the expression of BDNF gene is raised or decreased in a certain brain area, and the effect and mechanism of changes on the body;
(2) To identify changes in the epigenetic modification of BDNF in pain, and to observe changes in the DNA, histones, and miRNA of the BDNF gene from the molecular level in the brain region where BDNF expression has changed, in order to understand the effects of these changes on the BDNF gene;
(3) To clarify the mechanism of epigenetic modification of the BDNF gene in the body under chronic pain, and provide new ideas for the treatment of chronic pain. That is, blocking the pathways or molecules that affect the epigenetic regulation of the BDNF to inhibit changes in the epigenetic modification of the BDNF gene;
(4) Identification of targets for drug treatment of related diseases to prevent and treat the occurrence of multiple chronic pain. That is, the use of epigenetically-modified enzyme inhibitors to reverse the changes of the BDNF gene, normalizing the expression of the BDNF gene, thus achieving the purpose of preventing or improving the disease.