
Effect of Oral Honey Consumption on Immune Function Following Ex-Vivo Lipopolysaccharide Stimulation in Healthy Humans
Abstract
Nine healthy individuals with a mean ± SD BMI of 22.0 ± 0.7 kg/m² and age of 20 ± 0.2 years, participated in this single-blind randomised, crossover trial investigating the impact of ingesting two different honeys (1) Tropical Forest Honey (TFH) and (2) Manuka Honey; strength 12+ (MAN) on circulating levels of plasma interferon gamma following ex-vivo lipopolysaccharide (LPS) stimulation. Blood samples were prepared into duplicate aliquots of whole blood (800 μl) and 100 μg/l of LPS was added to samples to give a final volume of 1 ml. Levels of IFN-γ in plasma fractions were measured via commercially available sandwich ELISA and all comparisons were made with paired data using the Wilcoxon Signed Rank test taking a significance level of 5%.
Whilst significant intra-and-interpersonal variation was observed, IFN-γ concentrations remained statistically unchanged 48 hours after the ingestion of either honey (p=0.15). Thus, in this instance the type of honey did not influence the IFN-γ response to plasma samples spiked with LPS.
Article Information
- Received
- Accepted
- Published
Academic Editor: Sadanand Naik, HOD (Clinical Biochemistry), K.E.M. Hospital, Pune, India.
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2019 Olivia McCarthy, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Corresponding author: Olivia McCarthy, Applied Sport, Technology, Exercise and Medicine Research Centre (A-STEM), College of Engineering, Swansea University, Swansea, UK, Address: Swansea University, Bay campus, college of engineering, Fabian Way, Swansea SA1 8EN —
Competing Interests
The authors have declared that no competing interests exist.
Funding
No funding to report.
Data Availability
No data-availability statement was provided by the authors.
Acknowledgements
Investigation
Olivia McCarthy, Gareth Dunseath
Validation
Gareth Dunseath, Richard. M. Bracken
Writing – Original Draft
Olivia McCarthy
Writing – Review & Editing
Olivia McCarthy, Richard. M. Bracken, Othmar Moser, Max
L. Eckstein, Rachel Deere, Othmar Moser, Steve Bain, Gareth Dunseath
Statistics
Olivia McCarthy, Owen Bodger
With thanks to Robert Nash and all participants for their contributions to the study.
Disclosure Statement
O. McCarthy has received a Zienkiewcz PhD research scholarship. O. Moser has received lecture fees from Medtronic, travel grants from Novo Nordisk A/S and EASD and research grants from Sêr Cymru II COFUND fellowship/European Union, Novo Nordisk A/S, Novo Nordisk Austria and Dexcom. M. L. Eckstein has received a KESS2/European Social Fund scholarship. S. C. Bain reports having received honoraria, teaching and research grants from the Abbott, Astra Zeneca, Boehringer Ingelheim, BMS, Diartis, Eli Lily and Company, GlaxoSmithKline, Johnson & Johnson, Merck Sharp & Dohme, Novartis, Novo Nordisk, Pfizer, Roche, Sanofi-Aventis, Schering-Plough, Servier and Takeda. R. M. Bracken reports having received honoraria, travel and educational grant support from, Beneo, Boehringer-Ingelheim, Eli Lily and Company, Novo Nordisk, Sanofi-Aventis. R. Deere has no disclosures to report. This work was not carried out in the presence of any personal, professional or financial relationships that could potentially be construed as a conflict of interest.
Funding
No funding to report.
Citation:
Introduction
As the body’s largest organ of host defence the vitality of the microflora that reside within the gastrointestinal tract is critical in minimising pathophysiological complications1. The functionality of the microfloria is largely influenced by variations in microbial communities. These communtities are shaped by a series of complex and dynamic epignetic factors including diet, lifestyle and disease2. Interactions between the gut microbiota and the host immune system can modulate metabolic reactions as exemplified by the production of bile acids, choline and short chain fatty acids that are essential for host health2. This reinforces the nexus between host nutritional status, the microbiota/microbiome and reactionary immune responses. Appreciating these axes is a prerequisite for optimising therapeutic strategies that have the potential to elicit a protective immune response. With continuing financial strain on our national health services and the heavy reliance on pharmaceutical drugs as counter measures to fight disease, the discovery of dietary stratergies that target the prevention rather than cure manifesto are worthy of further exploration.
Honey, is a viscous and hygroscopic substance produced by bees from floral nectar. This sugar-rich liquid is pivotal for plant ecology, insect evolution and biodiversity. Historically, honey has been used as a natural healing agent, with illustrative evidence of its use dating back some 8000 years. Conventionally, honey is recognised as a natural therapeutic agent due to it’s incredibly nutrient dense profile3. Almost all ‘raw’ (i.e. unpasterised) honey contains several micronutrients (calcium, iron, magnesium, phospohorous, potassium, sodium and zinc), vitamins (B2, B3, B5, B6, B9 and C) and enzymes (glucose oxidase), which proposedly contribute to the potent inhibitory effect it evokes on up to 60 species of bacteria, fungi and viruses4. Whilst composed primarily of fructose and glucose, honey also contains the prebiotic fructo-oligosaccharide (FOS). Once fermented in the colon, FOS stimulates the production of bifidobacteria, one of the major genera of bacteria that characterise a ‘healthy’ microflora.
Unsurprisingly, the multifunctional immunomodulatory properties of honey have attracted scientific attention, with research referencing it’s antimicrobial5, anti-inflammatory6, antioxidant7, and anti-mutagenic properties6,8. Of particular interest are the immunological properties of honey, which depending on it’s biogeographical source can contain elevated levels of naturally-occurring casuarine glucoside; a compound associated with actively priming a stronger pro-inflammatory response to pathogens. Importantly, casuarine glucoside is an upstream regulator of various pleiotropic cytokines including interferon gamma (IFN-γ), a dimerised, soluble cytokine which enhances cell-mediated immunity and activates natural killer (NK) cells to provide antiviral activity9.
Clinically speaking, IFN-γ represents a crucial mediator of several cellular processes that help alleviate various pathologies e.g. multiple sclerosis10, organ transplantation11 and rheumatoid arthritis12. IFN-γ is also an important indicator of the activation of delayed type hypersensitivity T-cells (TDTH), which release IFN-γ as a “recall” response to antigens. Although mostly regarded as a T-cell associated cytokine, IFN-γ is also produced by other immune cells13. Indeed, combined stimulation with both IFN-γ and lipopolysaccharides (LPS) or LPS alone can significantly increase the percentage of IFN-γ producing monocytes in healthy individuals13. LPS/endotoxin is a cell wall constituent of gram-negative bacteria that activates macrophage microbicidal effector functions as well as the production of various pro-inflammatory cytokines. Upon entry into the circulation, LPS stimulates an acute phase anti-inflammatory response via the upregulation of macrophages which attempt to fight pathogenic invasions.
Interestingly, recet pilot work by Davidson and Rotondo (2005) observed the potent anti-inflammatory effects of orally consuming 20.4g (1.3 mg casuarine glucoside concentration) of Tropical forest honey (TFH) at 0, 2, 7, 14 and 21 days after administration. Peak increases in IFN-γ were found on day 2 i.e. 48 hours post consumption, with values reaching 4.6, 5.0, 9.7 and 13.0-fold higher than baseline levels (day 0) in response to 0 μg/ml, 0.01 μg/ml, 1 μg/ml and 100 μg/ml lipolysacharide (LPS) respectively.
Considering that even small amounts of IFN-γ could enhance the ability of macrophages to fight pathogens14, the elevated concentrations observed during this pilot study are noteworthy. However, although generally considered a nutrient dense food, substantial inter-and-intra-variations in both the macro-and-micronutrient profile of any given honey can exist15. Such differentiations are largely dependent on variations in its complex chemical composition which can vary considerably depending on nectar origin, bee species, geographical location and production procedure. Therefore, comparative work that investigates potential differences in immunological biomarkers between honey varieties may assist the identification of those most suited to pathophysiological complications.
The aim of this study was to observe the impact of orally ingesting two different honeys (1) Tropical Forest Honey (TFH) vs (2) Manuka Honey; strength 12+ (MAN) on circulating levels of plasma IFN-γ following ex-vivo lipopolysaccharide (LPS) stimulation in healthy humans.
Materials and Methods
Participants and Background Methods
Following approval by a local university ethics committee, nine healthy individuals were included in this trial with a (mean ± standard deviation (SD)) BMI of 22.0 ± 0.7 kg/m² and age of 20 ± 0.2 years. Due to the exploratory pilot nature of this work, a small participant number was deemed acceptable. Following familiarisation sessions, participants were selected after the completion of a health questionnaire confirming they were free from any underlying health issues and consented prior to commencing experimental trials.
The study used a single blind- crossover design including a 4-week wash out phase having participants randomised to orally consume 20.0 ± 0.02g of either Tropical Forest Products Zambian honey (TFH) or Manuka honey+12 (MAN). Each participant undertook 4 visits in total. The primary outcome of the study was to observe any differences in INF-γ concentrations following ex-vivo LPS stimulation after orally consuming the two honeys.
Experimental Day Procedures
Following a 2-day dietary recall, fasted-baseline venous blood samples (6 ml) were taken immediately before and 48 hours after the oral consumption of 20.0 ± 0.02g of randomly allocated honey. This procedure was repeated after a 28-day wash-out period when participants returned to consume the alternative honey. The wash out period reflected previous pilot research which demonstrated a peak increase in plasma IFN-γ concentration in response to 100 pg/ml PPD 48 hours post consumption, which decreased in a time dependant manner up to 21 days’ post ingestion16. On each visit, participants were taken through anthropometric measurements and completed a 48-hour dietary recall having only consumed water ad libitum in the 12 hours prior to each visit.
Blood Preparation Procedure
Duplicate aliquots of whole blood (800 μl) were prepared under aseptic conditions into sterile lithium heparinised 1.5ml vials. LPS (Sigma Chemicals), at a concentration of 100 μg per ml were added to the samples to give a final volume of 1ml. The samples were incubated for 20 hours at 37°C, 5% CO2, 100% humidity. At the end of the incubation period the tubes were centrifuged at 14.000 xg: 30 seconds, the plasma was decanted and stored at –20˚C until assayed for human IFN-γ according to manufactures instructions (DIF50 Human IFN-γ Quantikine ELISA Kit; R & D systems). Intra-assay and inter-assay variation were ≤4.7% and ≤7.8% respectively. Intra-assay variation was calculated by running 20 replicates from 3 individual samples in a single assay while the inter-assay variation was from 3 individual samples run in 40 separate assays.
Statistical Analysis
Plasma IFN-γ (pmol/l-1) concentrations were calculated using a curve fit software (MikroWin). The average of the duplicate readings for each standard, control, and sample, was taken and the average zero standard optical density was subtracted. Data are reported as mean ± SD. The data consisted of before and after values for each participant at both phases of the study. All variables showed moderate to severe positive skew and, given the extremely small sample size, could not easily be transformed into a distribution compatible with the use of parametric methods. It was therefore appropriate to apply non-parametric methods. Since the study was constructed as a crossover study all comparisons can be made with paired data using the Wilcoxon Signed Rank test (WSR) taking a significance level of 5%.
Results
Grouped Mean Responses
Although in neither case was the result significant, absolute IFN- γ concentrations fell from baseline for both treatments (Table 1). Observationally, the degree of reduction was more pronounced following the consumption of TFH honey, whilst levels scarcely fell following Manuka (TFH; -284.69 ± 478.04 (-41%) versus 75.42 ± 246.95 (-21%)). However, in neither case was the result significant (p=0.139 and p=0.859 respectively; WSR).
Table 1. Summary table of IFN-γ concentrations (pmol/l-1) in each condition and time point| Baseline TFH | 48 hr post TFH | ∆ TFH | Baseline Manuka | 48 hr post Manuka | ∆ Manuka | |
| Mean | 694 | 409 | -285 | 359 | 284 | -75 |
| Minimum | 189 | 136 | -256 | 0 | 2 | -93 |
| Maximum | 1887 | 1065 | 1279 | 1572 | 869 | 703 |
| Median | 465 | 264 | 101 | 169 | 168 | -2 |
| IQR | 217, 1258 | 190, 665 | -33, 588 | 87, 509 | 127, 446 | -71, 95 |
| p-value | - | - | 0.139 | - | - | 0.859 |
Individual Responses
Although grouped data exemplified a drop-in plasma IFN-γ concentrations regardless of the type of honey consumed, findings of a count response using individualised data revealed that 5/9 participants (56%) reported an increase in absolute plasma IFN-γ concentrations following MAN consumption, whilst only 3/9 (33%) reported increases following TFH consumption (Figure 1. a). Moreover, when the data was relativised as a percentage change from baseline, the magnitude of the decrease following TFH appeared to be greater than the drops experienced following MAN (-21.80% versus -15% respectively). However, whilst further analysis comparing Manuka directly with TFH found some evidence of a difference (p=0.110; WSR) this fell below the threshold for significance. (Figure 1. b)
Figure 1. illustrates the individualised (a) absolute (pmmol/L-1) and (b) relativised percentage (%) change in plasma IFN-γ concentrations following LPS stimulation at baseline and 48 hours following the consumption of each honey; Tropical Forest Honey (TFH) versus Manuka Honey (MAN).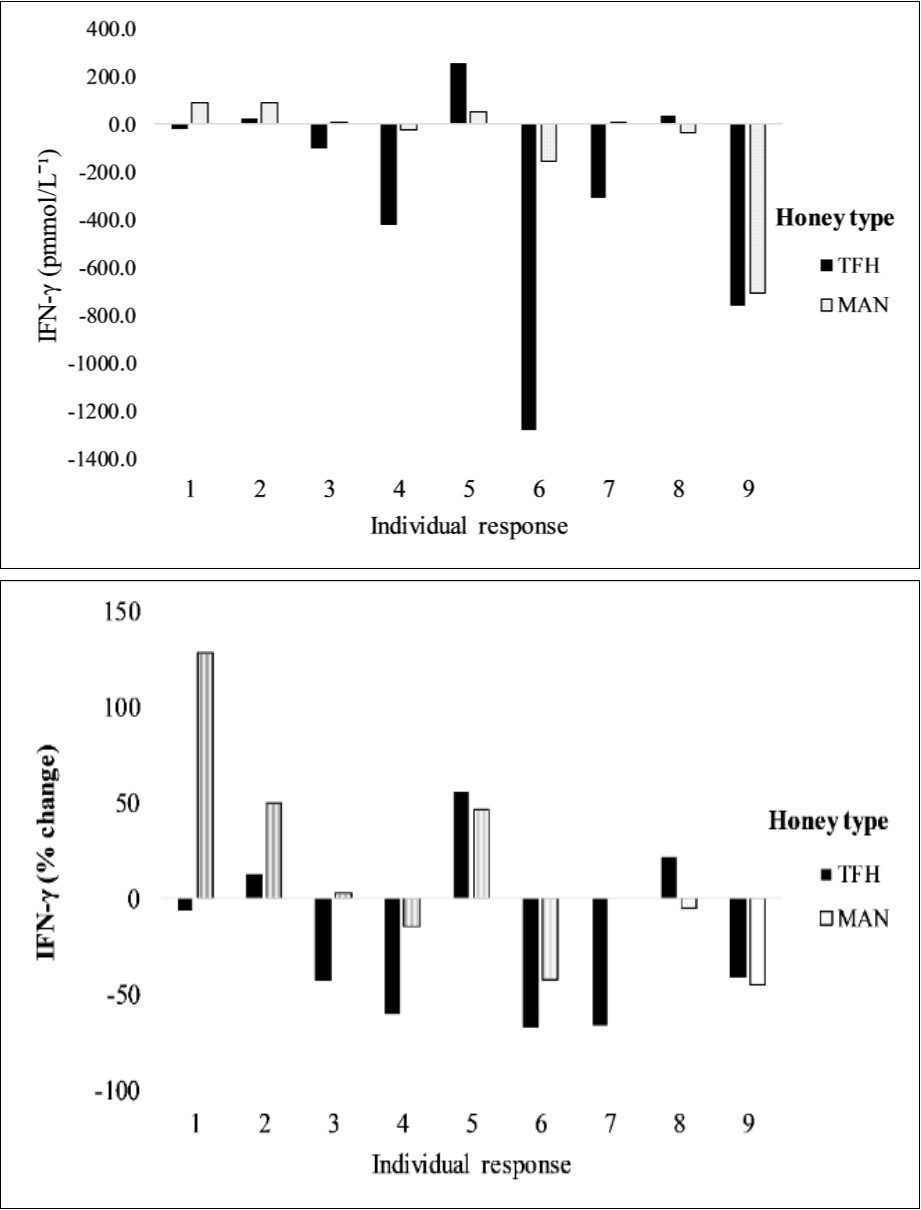
Download figure
Finally, it should be noted that comparative analysis found considerable difference in baseline plasma IFN-γ (pmol/l-1) concentrations under both experimental treatments (p=0.051; WSR). There was weak evidence (p=0.086; WSR) that IFN-γ levels were elevated before the second treatment, relative to those taken before the first treatment (although the ordering of treatments varied between participants) (Figure 2).
Figure 2. Change of plasma IFN-γ concentrations following LPS stimulation before and after oral honey consumption; Tropical Forest Honey (TFH) versus Manuka Honey (MAN).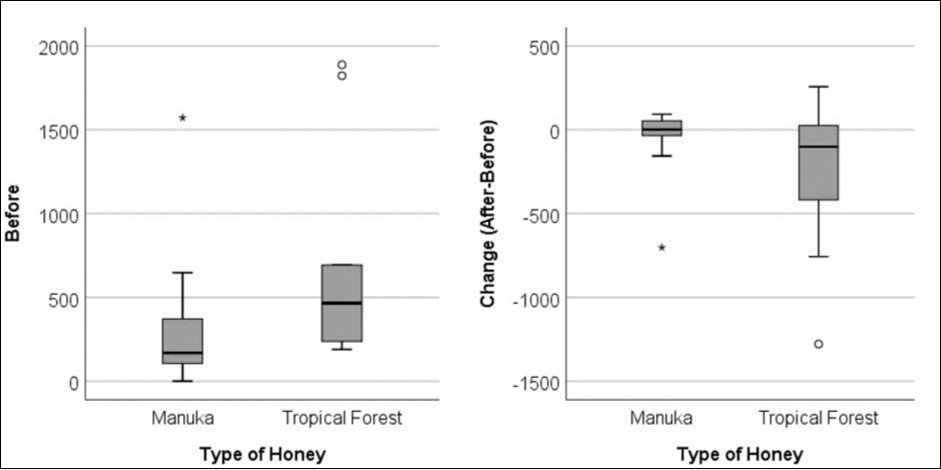
Download figure
Discussion
The aim of this study was to observe the impact of orally ingesting two different honeys (1) Tropical Forest Honey (TFH) vs (2) Manuka Honey; strength 12+ (MAN) on circulating levels of plasma IFN-γ following ex-vivo lipopolysaccharide (LPS) stimulation in healthy humans.
The results of this study demonstrate that although there was a natural pro-inflammatory response when plasma samples were spiked with 100 μg/ml LPS, this response remained unchanged 48 hours following the oral consumption of either TFH or MAN honey. Thus, in this instance, there was no discernible difference between the honeys on IFN-γ concentrations following ex vivo LPS stimulation.
However, it should be noted that whilst outside of statistical significance, a greater reduction in IFN-γ concentrations was observed 48 hours following the consumption of TFH versus Manuka honey (p=0.139 and p=0.859 respectively; WSR). Furthermore, when MAN and TFH were compared directly, there was some (albeit outside of the threshold statistical significance) evidence of a difference in the IFN-γ response (p=0.110; WSR). Moreover, findings of a count response revealed 5/9 (56%) participants reported an increase in absolute plasma INF-γ concentrations following MAN consumption, whilst only 3/9 (33%) reported increases following TFH consumption (Figure 1. a). Moreover, when the data was relativised as a percentage change from baseline, the magnitude of the drops following the consumption TFH appeared to be greater than the drops experienced following MAN (-21.80% versus -15% respectively). Collectively, these data may suggest potential differences in the anti-inflammatory properties of TFH and MAN honey, thereby agrees with the findings of previous literature which identifies variations in the biochemical constituents of different honeys. The clinical utility of identifying potential variations in the biological response to different honeys is of particular importance for selecting the appropriate choice for treatment purposes.
Conclusion
Whilst it transpires that there is a natural upregulation in the production of IFN-γ when following ex-vivo LPS stimulation, the magnitude of this response following the consumption of two different types of honey remained unchanged.
Despite statistical insignificance, the results from this study suggest the potential for differences in the biological response to different honey types, thus encourages the need for further research to explore dietary strategies that may elicit immunological properties.
The results from this pilot study also highlight the significant variation between individual responses (Figure 1 a and b) regardless of any attempts made to control for extraneous variables in the 24 hours prior to each experimental visit i.e. refrainment from exercise, alcohol and alterations in dietary intakes. Future research would benefit from exploring the effect of manipulations to the dosing frequency and quantity as well as employing an aged and/or patient population cohort who possess an inferior T-helper-1 response.
References
- 1.Ouwehand A, Isolauri E, Salminen S. (2002) The role of the intestinal microflora for the development of the immune system in early childhood. , Eur J Nutr 41(0), 1-1.
- 2.Nicholson J K, Holmes E, Kinross J. (2012) . , Host-Gut Microbiota Metabolic Interactions. Science(80-) 336(6086), 1262-1267.
- 3.Eteraf-Oskouei T, Najafi M. (2013) Traditional and modern uses of natural honey in human diseases, a review. , Iran J Basic Med Sci,Accessed,December2,2017,http://www.ncbi.nlm.nih.gov/pubmed/23997898 16(6), 731-742.
- 4.Eteraf-Oskouei T, Najafi M. (2013) Traditional and modern uses of natural honey in human diseases, a review. , Iran J Basic Med Sci,Accessed March5,2019,http://www.ncbi.nlm.nih.gov/pubmed/23997898 16(6), 731-742.
- 5.Tonks A J, Dudley E, Porter N G. (2007) A 5.8-kDa component of manuka honey stimulates immune cells via TLR4. , J Leukoc Biol 82(5), 1147-1155.
- 6.Othman N H, Hayati N. (2012) Honey and Cancer: Sustainable Inverse Relationship Particularly for Developing Nations—A Review. Evidence-Based Complement Altern Med. 1-10.
- 7.Erejuwa O O, Sulaiman S A. (2012) . , Ab Wahab MS. Honey: A Novel Antioxidant. Molecules 17(12), 4400-4423.
- 8.Wang X-H, Andrae L, Engeseth N J. (2002) Antimutagenic effect of various honeys and sugars against. http://www.ncbi.nlm.nih.gov/pubmed/12405798.Accessed June14,2017 , Trp-p-1. J Agric Food Chem 50(23), 6923-6928.
- 9.Gleeson M, Bishop N, Walsh N. (2017) . Exercise Immunology, https://www.amazon.co.uk/ Exercise-Immunology-Michael-Gleeson-ebook/dp/B00DP3JJJM,Accessed June,14
- 10.Huseby E S, Liggitt D, Brabb T, Schnabel B, Ohlén C et al.A pathogenic role for myelin-specific CD8(+) T cells in a model for multiple sclerosis. http://www.ncbi.nlm.nih.gov/pubmed/11535634, Accessed October2,2018 , J Exp Med 194(5), 669-676.
- 11.Xie L, Ichimaru N, Morita M.Identification of a novel biomarker gene set with sensitivity and specificity for distinguishing between allograft rejection and tolerance. , Liver Transpl 18(4), 444-454.
- 12.Cooper S M, Sriram S, Ranges G E.Suppression of murine collagen-induced arthritis with monoclonal anti-Ia antibodies and augmentation with IFN-gamma. October2,2018, http://www.ncbi.nlm.nih.gov/pubmed/3139747 , J Immunol 141(6), 1958-1962.
- 13.Kraaij M D, EJF Vereyken, PJM Leenen. (2014) Human monocytes produce interferon-gamma upon stimulation with LPS. , Cytokine 67(1), 7-12.
- 14.Schroder K, Hertzog P J, Ravasi T, Hume D A. (2004) Interferon-γ: an overview of signals, mechanisms and functions. , J Leukoc Biol 75(2), 163-189.
Cited by (1)
This article has been cited by 1 scholarly work according to:
Citing Articles:
International Journal of Complementary & Alternative Medicine (2023) OpenAlex