
Microanatomy of Thalamic Radiations
Abstract
Background
Thalamic radiations also known as thalamocortical pathways are reciprocal myelinated nerve fibers, arranged in a fanning pattern, grouped into tracts or fasciculi; and connecting the thalamus to the cerebral cortex. Detailed in vitro study of these tracts is seldom reported in the literature.
Objective
We sought to describe the microanatomy of thalamic radiations by means of the fiber-dissection technique to discuss challenges in dissection techniques and anatomic nomenclature, and follow through with a literature review.
Methods
Twenty formalin-fixed normal human hemispheres were dissected according to Klingler’s fiber-dissection technique under operative microscope.
Results
Thalamic radiations are reciprocal myelinated nerve fibers connecting the thalamus to the cerebral cortex and are referred to as corticothalamic and thalamocortical tracts. They are the most medial fibers of the internal capsule and consist of anterior (thalamofrontal), superior (thalamo-fronto-parietal or thalamoparietal), posterior (thalamooccipital) and inferior (thalamotemporal) thalamic fasciculi.
Conclusion
From the cerebral cortex, thalamic radiation fibers fan out into the thalamus and are the most medial fibers of the internal capsule. There is a great deal of controversy surrounding the distinction between anterior and superior thalamic radiations, sub-ependymal stratum and the fronto-occipital fasciculus.
Author Contributions
Academic Editor: Hesham N. Mustafa, Associate Professor of Anatomy, Faculty of Medicine, King Abdulaziz University, Jeddah, Saudi Arabia
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2017 Medard kakou, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Competing interests
The authors have declared that no competing interests exist.
Citation:
Introduction
Detailed information regarding in vitro study of thalamic radiations is seldom found in the extant medical literature. Déjerine9 (1895), Crosby7 (1962) and Nieuwenhyus20 (1988) successively demonstrated the existence of a superior occipital bundle. In 1909, Curran described for the first time the occipitofrontal fasciculus without mention of the superior occipitofrontal fasciculus. Tϋre demonstrated through his seminal work on brain dissection the absence of a superior occipital fasciculus28. Recent advances in diffusion-tensor MRI study of the brain white matter have shown the existence of other bundles such as an association tract connecting Broca and Wernicke areas, other than the superior longitudinal fasciculus4 and the inferior occipitofrontal fasciculus3, 19, 30. It would be interested to investigate the inferior occipitofrontal bundle using the Klingler dissection technique. We sought to describe the microanatomy of thalamic radiations by means of the fiber-dissection technique to discuss challenges in dissection techniques and anatomic nomenclature, and follow through with a literature review
Materials and Methods
Anatomic Specimen Preparation
Twenty normal human brain hemispheres were dissected according to Klingler’s fiber-dissection technique18regardless of gender. Specimens were taken from adult cadavers with no intracranial pathology nor history of such pathology in their medical history. Brain hemispheres were removed 36 hours after death and immediately immersed in water, then fixed in a 5% formalin solution in which specimens were suspended by a string attached to the basilar artery. This helped preserve the morphology of specimens by restricting their contact with the bottom of the crystallizer. Formalin solutions were renewed on a weekly basis for 3 months until the brain parenchyma became firm but not hard. After cleaning with running water, specimens were frozen at -15°C for 3 to 5 days. Afterwards they were allowed to thaw in water at room temperature, then dissected and stored back into 5% formalin solution. The effect of this process is to loosen up gray matter so that individual white matter nerve bundles delineate clearly18.
The cortex was resected using curets and underlying white matter fasciculi could be followed progressively by peeling them off under the operating microscope (Zeiss OPMI 9FC Oberkechen Germany). Stepwise dissection of fasciculi was documented with a digital camera and 2D images were then obtained.
Dissection Technique
Dissections started with the identification of a region-of-interest on a coronal view of a cerebral hemisphere using both diffusion-tensor magnetic resonance imaging tractography and data from anatomic literature. Lateral and medial surfaces of cerebral hemispheres were carefully dissected. During the dissection, slices were made whenever identification of new target regions proved necessary. Theoretically the superior occipitofrontal fasciculus (SOFF) should connect the frontal and occipital lobes through the parietal white matter lateral to the caudate nucleus6, 19, 20. But according to the results of the diffusion-tensor MRI3, 19, 30, the bundle of fibers uniting the frontal and parietal lobes could be referred to as the SOFF.
The stepwise dissection started from the lateral surface of the cerebral hemisphere, then moved to the medial surface using diffusion-tensor MRI tractography as a dissection guide.
At the lateral surface of the cerebral hemisphere, with or without operative microscope, we resected the cortex from the top of gyri to the deepest sulci using curets and wooden blunt-ended spatulas. As a result, U-shaped or arcuate association fibers connecting adjacent gyri were exposed. The gradual zabscission of these fibers from frontal, parietal and temporal opercula exposed the insula which cortex was ablated to discover the extreme capsule.
At the medial surface of the cerebral hemisphere, we resected the rostrum, genu of the corpus callosum and the cingulum respectively. The excision of the ependyma allowed us to expose the thalamus and the head of the caudate nucleus which, when ablated led us to the fibers of the superior occipitofrontal fasciculus as described by the diffusion-tensor MRI. The SOFF was removed completely to expose, in front, the thalamic radiations which are the most medial fibers of the internal capsule, and behind, the tapetum of the corpus callosum which is a sub-ependymal structure.
Results
Thalamic radiations or thalamocortical pathways are reciprocal myelinated nerve fibers, arranged in a fanning pattern, grouped into tracts or fasciculi; and connecting the thalamus to the cerebral cortex (Figure 2, Figure 3, Figure 4). They are the most medial fibers of the internal capsule and consist of anterior (thalamofrontal), superior (thalamo-fronto-parietal or thalamoparietal), posterior (thalamooccipital) and inferior (thalamotemporal) thalamic fasciculi.
Figure 1.(A) Coronal view of the left hemisphere and (B) global view after magnification and before dissection of (A). 1. Thalamus, 6. Lateral ventricule 7. Subependymal substratum, 9. Corpus callosum, 11. Internal capsule, 12. External capsule, 13. Claustrum, 14. Extreme capsule, 16. Upper branch of the SLF , 20. Head of caudeus nucleus, 23. Putamen, 24. Globus pallidus, 25. Lower branch of SLF 28. Insula, 30. Corona radiata, 32. Parietal pole, 33. Temporal pole.
Anterior and superior thalamocortical pathways course obliquely and connect anterior, mediodorsal, lateral ventral anterior, lateral dorsal and lateral posterior thalamic nuclei to the cerebral cortex. They exposed after the removal of the head and body of the corpus callosum were . A coronal view of the thalamus confirms the course of these fibers to the thalamus. The sub-ependymal stratum and gray matter are medial to the thalamic radiations (Figure 1). The successive excision of the insular cortex, claustrum, fibers of the extreme capsule, the putamen and globus pallidus and pyramidal tract exposed other components lateral to the internal capsule (Figure 1, Figure 4). Fibers of the extreme and external capsules and of the lenticular nucleus (putamen and globus pallidus) formed an inferior relationship with the stem of the uncinate and the inferior occipitofrontal fasciculi. A coronal section through the cerebral hemisphere and the thalamus completed the study of thalamic radiations. The optic radiations and the ventral amygdaloid tract which loop through the anterior perforated substance and the lateral expansion of the anterior commissure sheathed by the Gratiolet canal, which were not include intentionally in our illustrations, were further away. Gratiolet optic radiations connect the thalamus to the geniculate body and the posterior nucleus of the occipital cortex. These thalamic radiations line up the lateral ventricle.
The thalamus sends fibers in all directions. Anteriorly, the thalamo-fronto-parietal fasciculus connects the anterior and medial lateral nuclei to the cortex of the frontal pole. Superiorly, the thalamo-parietal bundle unites the anterior and intermediate latero-ventral nuclei to the posterior frontal and prefrontal cortex, and the lateral postero-ventral nuclei to the sensory associative parietal cortex. Posteriorly, the thalamo-occipital fasciculus connects the lateral geniculate body and the posterior nucleus to the occipital cortex. Posterior and superiorly, the thalamo-parietal fasciculus connects the dorso-lateral nuclei to the posterior associative cortex.
Posterior and inferiorly, the thalamotemporal fasciculus unites the posterior nucleus and the lateral geniculate body to the temporal cortex.
Thalamic Radiations Relationships with other Tracts
Our dissections failed to identify the occipitofrontal fasciculus connecting the frontal and occipital lobes which was different from the sub-ependymal stratum and anterior and superior thalamic radiations. The following is our stepwise dissection as anatomic structures are exposed in the order they appear. The removal of the external capsule and claustrum exposed the lateral surface of the putamen.
The internal capsule was exposed after the excision of the putamen and globus pallidus which were perfectly identifiable. The abscission of the outermost fibers of the internal capsule situated approximately 26 mm from the cerebral cortex and the head of the caudate nucleus exposed thalamic radiations (Figure 3).
The abscission of the frontal, parietal and temporal opercula exposed the superior longitudinal fasciculus (SLF) which was the first hemispheric and the largest association bundle to be seen and is located medial to the insular cortex. The SLF is an arcuate fasciculus which wraps around the insula and connects the frontal, parietal, occipital and temporal lobes together (Figure 1, Figure 3). It has a superior and an inferior surface, anterior or frontal end, posterior extremity and the lateral and medial margins. The lateral opercular border is flattened and more spread across. Its fibers project to the straight gyrus which was exposed after the withdrawal of the insular arcuate fibers. The medial edge is in contact with the corona radiata. The frontal, parietal and temporal opercula are lateral to the SLF. Ablation of the lenticular nucleus exposed the inferior and medial aspect of the SLF. Superiorly, U-shaped fibers are 13mm away from the cortex at the level of the central sulcus. The anterior extremity is lateral to the fanning end of the uncinate and inferior occipitofrontal fasciculi, then the corona radiata and the frontal horn of the lateral ventricle. This anterior end was lateral to the ventricle and 19,6mm away from the cortex (10-25mm). The posterior extremity was separated from the lateral ventricle by the optic radiations and the tapetum of the corpus callosum.
Figure 2.Subependymal stratum: (A) lateral view of right hemisphere and (B) medial view of left hemisphere. 1.Thalamus, 2. Anterior thalamic radiation, 7. Subependymal substratum, 4. Ependymal membrane, 6. Lateral ventricle, 11. Splenium of corpus callosum, 21. Cingulum, 30. Corona radiata, 32. Parietal pole, 33. Frontal pole, 35. Occipital pole, 36 tract optic
Figure 3.Thalamic radiations: (A) lateral view and (B) medial view. 1. Thalamus, 2. Anterior thalamic radiations, 3. Superior thalamic radiations, 4. Posterior thalamic radiations, 5. Inferior thalamic radiations, 6. Ventricular landmark, 10. Tapetum of corpus callosum, 11. Splenium of corpus callosum, 22. Ependymal membrane, 23 .Pons , 24. Mesencephalon, 17. Optic tract, 18. Uncinate fasciculus, 31. Occipital pole, 36. Cerebellum, 37.Frontal pole.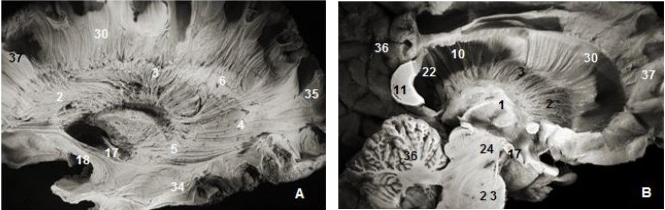
The C-shaped uncinate fasciculus (UF) connects the frontal and temporal lobes. The uncinate fasciculus (Figure 2, Figure 4) is composed of 3 parts: a fanning frontal end, an insular segment where the fibers are grouped together and a polar temporal extremity. The insular segment was discovered after the removal of the insular cortex laterally, the extreme capsule medially which consists of arcuate fibers connecting the insula to the opercula except the portion near the falciform fold. The last plane was exposed after the ablation of the claustrum and the extreme capsule which are below the tip of the insular cortex which is lateral to the limen insulae and medial to the claustrum. At this level, the posterior and superior margins were closely attached th lower edge of the inferior occipito frontal bundle. The detachment and excision of this fasciculus exposed the frontal and temporal rapports of the uncinate fasciculus.
The frontal end was discovered after complete removal of the orbital part of the lower frontal gyrus, the pars orbitalis of the upper frontal gyrus and the subcallosal area. The anterior and inferior surfaces were part of the internal capsule. Its anterior margin sweeps from below upward from the isthmus, the lateral edge of the anterior perforated substance and the junction between the lateral and inferior surfaces of the head of the caudate nucleus respectively. Its posterior edge was attached along its full length to the anterior border of the inferior occipitofrontal fasciculus (IOFF). The medial surface of the upper frontal end covered the anterior edge of the putamen and the inferior surface of the head of the caudate nucleus.
The temporal end is distributed to the uncus, the amygdala and the temporal gyri. Here too, the detachment and excision of the IOFF helped the study of the temporal distributions of the uncinate fasciculus. The insular segment was 2mm thick and 8.8mm wide.
Figure 4.(A) Lateral view of left cerebral hemisphere after dissection and (B) coronal view. 2. Anterior thalamic radiation, 7. Subependymal stratum, 8. Corpus callosum, 12. Internal capsule, 13. External capsule, 13. SLF, 15. Corona radiata, 16. Uncinate fasciculus, 19. Inferior frontal occipital fasciculus, 27. Putamen, 28. Globus pallidus, 29. lateral ventricle, 31. Parietal pole, 34. Temporal pole.
The inferior occipitofrontal fasciculus (Figure 2, Figure 4) closely attached to the previous fasciculus was composed of fiber bundles connecting the frontal lobe to the temporal, posterior parietal and occipital lobes. The IOFF is shaped somewhat like a flattened tape and is composed, as the uncinate fasciculus, of three parts: frontal, insular and parietal temporo-occipital segments.
The insular segment was fully exposed after the ablation of the uncinated bundle and courses medial to the external and extreme capsule and above the uncinate fasciculus. This segment was on average 2 mm thick and 8.8 mm wide.
The parietal temporo-occipital segment is about 20 mm wide stretching from the amygdala to the pulvinar and geniculate bodies at the height of the ventricular atrium.
The fanning frontal segment is located at the height of the frontal horn of the lateral ventricle. The frontal distribution of the inferior occipitofrontal fasciculus stretches to the whole of Broca's area. This frontal end was located 25 mm away from the cortex of the lower frontal gyrus. It is the second fasciculus connecting the frontal and the occipital lobes, and is thin at the frontal end.
The sub-ependymal stratum also known as the subcallosal stratum was made of a fiber-free layer of white matter. This layer of white matter was exposed after excision of the ependymal layer lining the ventricular system. It is located between the caudate nucleus and callosal radiations (Figure 1, Figure 2, Figure 4). Under the ependyma backward to the occipital horns, callosal fibers form together the tapetum and thalamic radiations. Our dissection failed to show a precise boundary between the tapetum and the sub-ependymal stratum. The substratum is a sub-ependymal structure located between the caudate nucleus, the corpus callosum radiations and the internal capsule. Globally, it is located below the ependyma and medial to the cingulum. It is not made of fibers connecting the frontal and the occipital lobes.
The Optic radiations, the ventral amygdaloid tract loop through the anterior perforated substance and the lateral expansion of the anterior commissure sheathed by the Gratiolet canal. Due to their remote location, we did not exposed them intentionally during our dissection. They connect the thalamus to the geniculate body and the posterior nucleus of the visual occipital cortex which is made of the optic radiations of Gratiolet .
Discussion
We agree with other authors about the fact that the superior occipitofrontal fasciculus is almost none existent.
The study of the superior occipitofrontal fasciculus faces technical challenge and anatomic nomenclature issues
Dissection Technique
The dissection of the cerebral white matter was abandoned in favour of microtome and histological studies. The arrival of the Klingler white matter dissection technique revolutionised cerebral white matter dissection as the best technique for white matter exploration, despite the fact that it is laborious and time consuming. It requires longer preparation time ranging from three months to a year. The longer the preparation time, the easier the dissection and the better the distinction between grey and white matter. Detailed information regarding the Klingler white matter technique is seldom found in the extant medical literature. Most publications concerning this dissection technique deal with anatomic regions of easy access like the temporal region and the lateral surface of the cerebral hemisphere8, 10, 13, 14, 15, 16, 17, 25, 26, 27, 28, 29. Klingler 18 and Ebeling are the only authors which works on the pyramidal tract11 stood out.
These technical challenges paved the way for the development in recent years of the diffusion-tensor MRI tractography which should be given credit as a new technology. This new technology was partially recognised in the seminal work of Kier on the uncinate fasciculus15. Questions remain on the credibility of his results given the fact that the dissection focused on a region of easy access and there are unanswered questions on the superposition in-vivo and ex-vivo tractography data.
The principle of tractography is to conduct a three-dimensional study of the heterogeneity of water molecules’ speed. The brain white matter is highly anisotropic14, 19, 22. Tractography is based on the measurement of the degree of anisotropy of white matter fibers which allow the study of their microstructural organization6, 14, 19, 22.
Our anatomic findings lead us to address common issues related to both methods. They both allow a three-dimensional study of white matter bundles. Tractography and the Klingler technique are based on the behaviour of water molecules between axons. In the Klingler dissection technique, ice formation between axons facilitate dissection, whilst the displacement or preferential orientation of water molecules in tractography facilitate fiber dissection of the cerebral white matter. In both techniques, crossing fibers are difficult to dissect1 and both are not appropriate in studying vascular relationship of neighbouring structures.
Nonetheless, the Klingler technique has the advantage of studying the cortex and basal ganglia grey matter, and it might help the understanding and validation of tractography. Tractography which needs improving, still faces issues of reproducibility and comparability of data and of correcting artefacts. This is true when analysing origins, junctions and the termination of fasciculi14. But tractography is non-invasive and can be done in-vivo or during surgery 1, 21.
Our anatomic region-of-interest identified as the superior occipitofrontal fasciculus presented dissection challenges in diffusion-tensor MRI tractography and in the Klinger technique alike, because it is located at the junction between callosal fibers and the internal capsule.
Difficulty of the Anatomic Nomenclature
Generally speaking, there is a great deal of controversy regarding the anatomic nomenclature of white matter fiber bundles in the extant medical literature probably due to the difficulty in their study. Déjerine (1895) mentioned the occipitofrontal fasciculus which matched the superior occipitofrontal fasciculus described by the diffusion-tensor MRI tractography3, 19, 30. He did not mention the superior occipitofrontal fasciculus and failed to distinguish the inferior occipitofrontal fasciculus from the uncinate fasciculus. Déjerine described it as Forel and Onufrwicz did, as being sagittal and medial to the corona radiata, above the caudate nucleus, below and lateral to the corpus callosum, and separated from the ventricle by the sub-ependymal gray matter. Meyert called this fasciculus the caudate nucleus corona radiata and Wernicke called it the corpus callosum fasciculus coursing the internal capsule. Déjerine reinforced his description and talked of a sagittal association fasciculus located medial to the corona radiata above the caudate nucleus, below and lateral to the corpus callosum; and it is separated from the ventricular cavity by the sub-ependymal grey matter. It is a long fasciculus located between the cingulum and the superior longitudinal fasciculus of Burdach 9. This fasciculus is separated from the cingulum by the entire thickness of the corpus callosum and the superior longitudinal fasciculus by the foot of the corona radiata. It looks like an open curve forward and downward, and it is similar to the one described by the diffusion-tensor MRI. In 1909 Curran described in detail the inferior occipitofrontal fasciculus and paradoxically did not mention its superior counterpart. In 1956 Klingler described a subcallosal fasciculus connecting the frontal and occipital lobes which is attached laterally to the caudate nucleus, and did not named it the superior occipitofrontal fasciculus 18. This fasciculus might had been described by Muratoff but there is no evidence of his work in the literature. Crosby (1962), in his treatise drew this fasciculus but failed to provide photographic evidence from his dissections 7. He described the superior occipitofrontal fasciculus as a bundle of white matter fibers connecting the occipital and temporal cortex to the frontal and insular cortex 7. Crosby and Nieuwenhus20 description of the superior occipitofrontal fasciculus is quite similar to that of the diffusion-tensor MRI. According to Türe the anterior and superior thalamic radiations and the sub-ependymal stratum should be referred to as the superior occipitofrontal fasciculus 28. He suggested that the inferior frontal fasciculus should be christened as the occipitofrontal fasciculus. Rhoton 24 in 2002 mentioned only the uncinate fasciculus and the inferior occipitofrontal fasciculus, and argued the presence of only one fasciculus. The superior part is made of fibers connecting the superior and lateral portions of the frontal lobe and the most lateral part of temporal gyri near the temporal pole, and is part of the uncinate fasciculus. It is therefore not the inferior occipitofrontal fasciculus.
As far as the superior longitudinal fasciculus is concerned, some author divide it into two fasciculi: the superior longitudinal fasciculus and the arcuate fasciculus 22. Others think, it is a single fasciculus with two branches, an anterior branch and a posterior branch18, 21, 29. Diffusion-tensor MRI identified a fronto-parietal fasciculus quite different from the superior occipitofrontal fasciculus3, 19, 30. But that description is still close to that of Crosby7 and Nieuwenhuys 20. The superior occipitofrontal fasciculus is located under callosal radiations, hence the name subcallosal fasciculus3; and between the cingulum and the superior longitudinal fasciculus9. Tractography data of this fasciculus showed a fasciculus different from the superior occipitofrontal fasciculus 3, 19, 30. These tractography data identified a fasciculus connecting only the frontal and the parietal lobes, which partially settle the question of anatomic nomenclature. In fact the region-of-interest is located medial to the junction of the internal capsule fibers and the callosal radiations and is difficult to dissect as we emphatically pointed out in our description of the Klingler technique.
Anterior and superior thalamic radiations whose fibers are the most medial, are closely attached to the sub-ependymal stratum and course obliquely toward the thalamus. Therefore tractography data could produce many artefacts and cause confusion during their interpretation. But the findings of many authors reinforce the real existence of this fasciculus which is a fronto-parietal fasciculus not the superior occipitofrontal fasciculus. Tractography data failed to formally identify the superior occipitofrontal fasciculus. Furthermore it is not easy to draw a line between this fasciculus and the anterior and superior thalamic radiations, given the fact that its anterior (frontal) and posterior (parietal) ends might well correspond to the anterior and superior thalamic radiations. Its deep location near the thalamus and caudate nucleus make it hard to easily distinguish it from the sub-ependymal stratum. It is the authors’ view that the combination of anatomic dissection and neurostimulation explorations will provide additional and convincing evidence, provided that data interpretation is robust and reliable.
Conclusion
The superior occipitofrontal fasciculus may well exist but our dissection failed to ascertain its authenticity. The present work aims to draw the attention of anatomists and neuroradiologists to design more ambitious studies to settle this anatomic nomenclature. A good understanding of clinical correlation related to its lesions requires clear description of the morphology and topography of this fasciculusprovide additional and convincing evidence, provided that data interpretation is robust and reliable.
Acknowledgements
We would like to thank sincerely Daniel Bourry for the beautiful illustrations, and Gérald Deluermoz and Jean-Paul Da Silva for their help in preparing anatomic specimen.
References
- 1.Berman J I, Berger M S, Mukherjee P, Henry R. (2004) Diffusion-tensor imaging – guided tracking of fibers of the pyramidal tract combined with intra operative cortical stimulation mapping in patients with gliomas. , J. Neurosurg 101, 66-72.
- 3.Catani M, Howard R J, Pajevil S, Jones.. DK (2002):Virtual in vivo interractive Dissection of White Matter fasciculi in the Human Brain .Neuroimage.17: 77-94.
- 4.Catani M, Jones K.FFytche F:Perisylvian(2005). , Language Networks of the Human Brain .Annneurol 57, 8-16.
- 5.Coppens J R, Mahaney K B.Abulrauf SI (2005): An anteromedial approach to the temporal horn to the optic radiation fibers and uncinate fasciculus:anatomical and technical note. Neurosurg focus. 18(6B): E3.
- 6.Cottier J P.barantin L,destrieux C wang R et al (2004) .Imagerie cérébrale en tenseur de diffusion et tractographie de la substance blanche :principes et limites actuelles Feuillets de Radiologie;. 45, 191-199.
- 7.Crosby E C.humphrey t and Lauwer EW (1962).Correlative anatomy of the nervous system .Mac millian co.NewYork.
- 8.Curran E. (1909) A new association fiber tract in the cerebrum. , J. Comp. Neurol. and Psycho.19: 645-657.
- 10.Ebeling U, Cramon D. (1992) Topography of the uncinate fascicule and adjacent temporal fiber tracts .Acta neurochirurgica (Wien). 115, 143-148.
- 11.Ebeling U, Reulen H. (1992) Sub cortical topography and proportions of the pyramidal tract. , Acta Neurochirg.(Wien).118: 164-171.
- 12.Ebeling U, Reulen H J. (1988) Neurosurgical topography of optic radiation in the Temporal lobe .Acta Neurochirurgica (Wien). 92, 29-36.
- 14.Habas C. (2004) Fondements physiques élémentaires de la tractographie en tenseur de diffusion. , J. Radiol.85 : 281-286.
- 15.Kier E L, Staib L H, Davis L M, Bronen R A. (2004) Anatomic dissection tractography. A new method for precise MR localisation of with matter tracts. , AJNR 25, 670-676.
- 16.Kier E L, Staib L H, Davis L M.Bronen RA (2004).MR imaging of the Temporal stem: Anatomic Dissection Tractography of the uncinate fasciculus, inferior occipito frontal fasciculus and Meyer’s loop of the optic. Radiation.ANJR.25: 677-691.
- 17.Peltier J, Travers N, Destrieux C, Velut S. (2006) Optic radiations: a microsurgical anatomical study.JNeurosurg.105(2):. 294-300.
- 18.Peltier J, Verclytte S, Delmaire C, Pruvo J P, Godefroy O.Le Gars D(2010). Microsurgical anatomy of the temporal stem: clinical relevance and correlations with diffusion tensor imaging fiber tracking.J. , Neurosurg 112(5), 1033-8.
- 19.Klingler J, Gloor P. (1960) The connections of the amygdala and the anterior temporal cortex in the human brain. , J. Comp. Neurol 155, 333-369.
- 21.Mori S, Kaufmannn W E, Davatzikos C, Stieljes B. (2002) Imaging cortical association tracts in the human brain using Diffusion Tensor based axonal tracking. Magnetic resonance in medicine.47;56:. 215-228.
- 22.Nieuwenhyus R, Voogd J, Van.Huijzen C (1988).The human central nervous system. Ed. third revised edition:Springer-Verlag.
- 23.Nimsky C, Ganslandt O, Hastreiter P, Wang R, Benner T et al. (2005) Preoperative and intraoperative Diffusion Tensor Imaging-based Fiber tracking in Glioma Surgery. , Neurosurgery 56, 131-138.
- 24.Oppenheim C, Rodrigo S, Poupon C, A Dumas de Roque, Naggara O et al. (2004) Imagerie en tenseur de diffusion et système nerveux central. Pour quelles applications cliniques ?. , J.Radiol; 85, 287-96.
- 25.Peuskens D, J Van Loon, F Van Calenbergh.et al (2004): Anatomy of the anterior temporal lobe and the fronto temporal region demonstrated by fiber dissection. , Neurosurgery 55, 1174-1184.
- 27.Sincoff E H, Tan Y, Abulrauf S I. (2004) White matter fiber dissection of the optic radiation of the temporal lobe and implication for surgical approaches to the temporal horn. , J. Neurosurg.101: 739-746.
- 28.Rubino P A, Rhoton AL Jr.Tong X Oliviera de e (2005). Three-dimensional relationships of the optic radiation. , Neurosurgery; 57(1), 219-225.
- 29.Türe U, Dianne C H, Yaşargil R N, Al-Mefty O, Yaşargil G. (1999) Topographic anatomy of the insular region. , J. Neurosurg 90, 720-733.
- 30.Türe U, Yaşargil G, Pait T G. (1997) Is there a superior occipitofrontal fasciculus?. , A Microsurgical Anatomic Study. Neurosurgery; 40, 1226-1232.
Cited by (10)
This article has been cited by 10 scholarly works according to:
Citing Articles:
NeuroImage Clinical (2025) OpenAlex
NeuroImage: Clinical (2025) Crossref
Montserrat Domingo-Ayllon, C. García-Gorro, Nadia Rodríguez-Dechicha, Irene Vaquer, Matilde Calopa et al. - NeuroImage: Clinical (2025) Semantic Scholar
European Journal of Neuroscience (2025) OpenAlex
S. Chester, J. C. Rogers, T. Ogawa, M. Terao, R. Nakai et al. - European Journal of Neuroscience (2025) Semantic Scholar
NeuroImage (2022) OpenAlex
NeuroImage (2022) Crossref
bioRxiv (Cold Spring Harbor Laboratory) (2021) OpenAlex
A. Radwan, S. Sunaert, K. Schilling, M. Descoteaux, B. Landman et al. - NeuroImage (2021) Semantic Scholar
Journal of Nihon University Medical Association (2021) OpenAlex
Journal of Nihon University Medical Association (2021) Crossref
K. Ogawa, Takayoshi Akimoto, Makoto Hara, M. Fujishiro, Yutaka Suzuki et al. - Journal of Nihon University Medical Association (2021) Semantic Scholar