Abstract
The management of symbiotic Microbial Biota (MB) in the soil as agents that promote the yield and health of crops, is aimed at inducing modifications of the phenotype of plants, both over and under the ground. It is here shown, in Sorghumsudanensis plants, that: i) a simple response to MB inoculation is the result of the fall out of the raw pH; ii) the simple NIR scans of leaves can be considered to rapidly classify the outcomes; iii) the raw pH can be considered a key-variable of leaf modifications. An experiment was carried out on Sorghumsudanensis. The plants were seeded in pots and grown for 66 d, and then a control non-inoculated group (C) was compared with thirteen Arbuscular Mycorrhizae (AM) Glomus inoculated groups and with two commercial MB products. A total of 374 raw pH measurements conducted on the leaves showed that the 5.18 pH units in the C group were scaled by -1.9% (P<0.0336) in the MB group and by -3.4% in the AM group (P<0.0001), with a relevant diversity between groups. Direct discrimination of these three groups, by means of smart NIR-SCIO, showed a % reclassification of the C, MB and AM groups of 74%, 59% and 96% in the fresh leaves and of 65%, 51% and 94% in the dried ground leaves, respectively. The composition of the dried leaves, based on a set of 14 variables predicted via NIRS models, plus the total foliar dry weight and percentage, showed a typical increase in protein, ash and hemicellulose, and a typical decrease in the cellulose, dry matter, crude fiber and crop maturity index. These variables were related to the foliar pH, as a key-variable, by means of a PLS standard model (R2 0.81) in which a low pH steadily favored the dry mass weight and, to a lesser extent, the hemicellulose and the digestible NDF contents; on the other hand, a high pH increased the dry matter percentage and the cellulose content of the leaves. As expected, the leaves of the inoculated plants showed a more juvenile ontogenic status. The epigean botanical modifications can be considered harmonic expressions of a luxuriant symbiosis, as testified by the homologous NIR categorization. The outlook for a symbiotic agriculture, with mycorrhizal plants, should consider the raw pH as a multifaceted variable.
Author Contributions
Academic Editor: Prem Narain, Biostatistics, Bioinformatics, Genetics Bachelor of Science (Hons), Master of Science, Doctor of Philosphy, Doctor of Science, USA.
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2018 Giorgio Masoero, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Competing interests
The authors have declared that no competing interests exist.
Citation:
Introduction
A credible increase of 50% in the world feed and food necessities can be expected at the end of the next thirty years1, and more fertilizers, water and pesticides will be needed to satisfy such needs. Starting all over again with a “Symbiotic agriculture” is a seductive “manifesto” for both a sustainable development of arable lands and for the new off-land agrotechnology2. Arbuscular Mycorrhizae (AM) need to be closely involved in a road map of sustainable agriculture as a result of their symbiotic capabilities in the agrarian multiverse, characterised by these highlights: 1) soil preservation, with restoration of the organic matter in both topsoil and polluted soils3; 2) anticipation of the Mitscherlich curve of phosphorus efficiency4; 3) resolution of the “nitrogen paradox” as a result of the uptake of organic-N 5; 4) benefits for the atmosphere and for the foreseen climate changes, with an important reduction in N2O emissions6; 5) luxuriance of the epigean as well as of the hypogean plant systems7; 6) enhancements of the functional properties of feeds and foods8, 9, 10, 11, 12 7) induction of greater intrinsic resistance to the onset of a pathological state, regardless of the origin and nature of the plants13.
Experiments with AM may be divided into two categories, that is, practical and scientific7. The first type analyses pure soils and substrates, such as those in a microbial desert, without agrarian biotas, while the second one studies extensive and multiple interactions in the presence of immense microbial charges14.
In a first field experiment with maize fertilized by a Micosat F® complex microbial consortium15, it was found that the in vivo raw pH of a plant was acidified according to a de-gradient from the roots (pH -7% in mycorrhizal maize) to the stem at an ear height (-4%). Other researches in vineyards confirmed the tendency of the raw pH16, measured at the petiole insert, to be lowered after microbial fertilisation with 10 kg ha-1 of MB compound inoculated near the secondary roots. The authenticity of the raw pH parameter was highlighted by Cornelissen et al.17, who considered the leaf pH as a new plant trait that could explain variations in the foliar chemistry and carbon cycling processes of subarctic plant species and types. The first work was followed by a microcosm experiment18, the results of which upset the myth of a pH dominance of the soil: the pH in fresh leaves will only grow for +0.036 pHsoil-1 (from -0.5 to -0.7%).
The aim of the present study has been to ascertain whether the decrease in the varied raw pH in leaves is due to AM or to a whole complex of a microbial consortium, and to assess how AM can be considered effective modifier agents of the Sorghumsudanensis phenotype, while considering the raw pH as a key-variable.
Experimental Procedure
Two rapid methods: the measurement of the raw pH, according to Masoero and Giovannetti15, and the smart NIR-SCIO measurement of the leaf, according to Masoero et al.19, have been used in this study to analyse the multifaceted effects of an inoculation of Sorghumsudanensis plants with thirteen AM Glomus strains and two microbial biotas fertilisers (MB), and the results have been compared with a non-inoculated control (C). Basically, we considered the in-vivo raw pH as a rapid indicator of symbiosis. In parallel, we considered the NIR spectra, reflected from the fresh and dried leaves (NIR-Tomoscopy), as a categorically discriminant tool. The third step concerned several traits of the leaf composition, which were predicted from an extended NIR-IR spectrum of the dried and ground tissues. Finally, the raw pH was connected, as a key-variable, to the phenotypes of plants by means of linear model analyses.
Materials and Methods
Plant material, Inocula and Growth conditions
Three categories were considered: i) Arbuscular Mycorrhizae (AM), with thirteen theses of crude inocula belonging to the Glomus genre from the USA (AM01-AM13); ii) Microbial Biota of the soil (MB), with two commercial approved bio-fertilizer consortia (MB1-MB2); iii) a non-inoculated Control thesis (C) (Table 1).
Table 1. Setup of the Categories and Theses.| Categories | Theses | Species |
|---|---|---|
| AM - Arbuscular Mycorrhizae | AM01 | Claroideoglomusetunicatum YV PA137 |
| AM - Arbuscular Mycorrhizae | AM02 | Paraglomusoccultum CL700A |
| AM - Arbuscular Mycorrhizae | AM03 | Acaulosporamorrowiae CL290 |
| AM - Arbuscular Mycorrhizae | AM04 | Gigaspora gigantea PA125 |
| AM - Arbuscular Mycorrhizae | AM05 | Sclerocystissinuosa MD126 |
| AM - Arbuscular Mycorrhizae | AM06 | Septoglomusconstrictum FL328 |
| AM - Arbuscular Mycorrhizae | AM07 | Gigasporarosea NY328A |
| AM - Arbuscular Mycorrhizae | AM08 | Funneliformusmosseae IT201 |
| AM - Arbuscular Mycorrhizae | AM09 | Rhizophagusintraradices CA502 |
| AM - Arbuscular Mycorrhizae | AM10 | Archaeosporatrappei FL208 |
| AM - Arbuscular Mycorrhizae | AM11 | Scutellospora pellucida MN408A |
| AM - Arbuscular Mycorrhizae | AM12 | Claroideoglomusclaoideum ON393 |
| AM - Arbuscular Mycorrhizae | AM13 | Diversisporaeburnea AZ420 |
| MB - Microbial Biota $ | MB1 | Micosat F© (CCS-Aosta, Quart, Italy) |
| MB - Microbial Biota £ | MB2 | MycUp© (Symborg, Murcia, Spain) |
| C- Control not inoculated | C |
Sorghumsudanensis seeds from cv “Puma” (Padana Sementi Elette S.r.l. Tombolo, Italy) were inoculated at seeding in 10 L pots (three seeds per pot), using 12 g pot-1 of crude inoculum. The growth substrate was volcanic lapillus rock (5 L) and peat (5 L). The substrate was kept in the greenhouse and drip irrigation was adopted.
The leaves were harvested after 66 d and measured to establish the raw pH of the leaves by means of direct contact of the split central vein with a combined plastic-glass Double Pore Knick® electrode (Hamilton, Reno, USA) using an “XSpH 70” pH meter (Giorgio Bormac S.r.l., Carpi, Italy).
The whole leaves from each of the 16 theses were chopped and dried at 60 °C to a constant weight, air-equilibrated, ground in a Cyclotec mill and stored for later analysis by means of NIRS.
Vibrational Spectroscopy and Chemometrics
The ground leaves were repeatedly examined (three scans) with a Spectrum IdentiCheckTM FT-NIR/MIR system (Perkin-Elmer, Beaconsfield, Buck, England) and the chemical composition was predicted via NIRS, using equations that were established on twelve different forage crops, as reported in Tassone et al.20.
NIR-Tomoscopy
A smart NIR miniaturized web-based wireless spectrophotometer, SCIO version 1.2 (Consumer Physics, Tel Aviv, Israel), with a 740-1070 nm range, was used to scan the fresh leaves on the upper foliar tinge (one scan) and the ground dry matrices (10 replicates).
Statistical Analyses
An ANOVA one-way model was used to fit the effect of the categories or of the 16 theses using PROC GLM from SAS-STAT 9.0 software (SAS Institute, Inc., Cary, NC, USA), while the Tukey test was used to adjust for LSMEANS PDIFF. Univariate analyses were conducted on the values of the raw pH and on the predicted composition.
The means values of the 14 variables pertaining to the 16 theses plus the total foliar dry weight and percentage, were then related to the foliar pH, as a key-variable, using a standard Partial Least Square (PLS) model (StatBox 6.5 v., Grimmer Logiciels, Paris, France) which allows two latent variables to be achieved; in this way, it was possible to identify the variables that were favoured or contrasted by the raw foliar pH variations among the theses.
A categorical discrimination of the three main groups of the fresh and dried leaves was performed by means of chemometrics of the 331-point spectra using the SCIO Lab proprietary software, based on AKA (Also Known As), the confusion matrix, without any mathematical pre-treatment of the spectra.
Results
Raw pH
The raw pH of the leaves appeared as a very narrow-distributed variable, which was characterized by a variation coefficient of 3.8% (Table 2).
Table 2. Raw pH values (No. 374) in the three Categories.| Statistics | Categories | Prob. | ||||
| C | MB | AM | Cvs. MB | Cvs. AM | MBvs. AM | |
| LSMeans | 5.18 | 5.10 | 5.00 | 0.0336 | <.0001 | 0.0002 |
| SE ± | 0.03 | 0.02 | 0.01 | |||
| RMSE | 0.19 | |||||
| R2 | 0.12 | |||||
The normal mean value of 5.18±0.03 in the leaves from the C non-inoculated plants was significantly decreased to 5.10±0.02 in the MB category (-1.5%, P=0.0336), and decreased to a value of 5.00±0.01 in the AM category (-3.5%, P<0.001). Moreover, the AM vs. MB difference was highly significant, with the MBs being less acidifying. When the individual theses (Table 3a and Table 3b; Figure 1) were considered, the differences appeared more scaled.
Table 3a. Results of the quality of Sorghum sudanensis leaves (No.48) £.| Thesis | pH | Dry Mass g | Hemicellulose % | NDF digestible % | Ether Extr. % | ADF, % | Crop Maturity Index | Crude Fiber % | Nitrogen Free Extract % |
| AM –Arbuscular Mycorrhizae | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| AM01 | 4.93b | 21.1 | 7.9 | 29.1 | 1.4 | 46.0 | 2.3 | 24.2 | 47.0 |
| AM02 | 4.96b | 14.3 | 7.1 | 28.9 | 1.7 | 45.5 | 2.1 | 24.6 | 42.5 |
| AM03 | 4.97b | 14.3 | 8.2 | 27.0 | 1.4 | 43.7 | 2.0 | 25.0 | 45.9 |
| AM04 | 5.03ab | 9.0 | 8.8 | 27.9 | 1.5 | 48.3 | 2.1 | 24.3 | 45.4 |
| AM05 | 4.95b | 16.0 | 6.1 | 27.4 | 1.7 | 48.8 | 2.4 | 26.5 | 42.7 |
| AM06 | 5.08ab | 17.2 | 6.4 | 27.3 | 1.5 | 45.8 | 2.1 | 24.0 | 45.6 |
| AM07 | 5.04ab | 16.9 | 4.8 | 27.7 | 1.4 | 46.9 | 2.2 | 23.9 | 43.9 |
| AM08 | 4.97b | 17.6 | 5.5 | 29.2 | 1.5 | 46.8 | 2.0 | 23.7 | 43.5 |
| AM09 | 5.03ab | 15.7 | 8.3 | 27.4 | 1.6 | 44.6 | 1.9 | 20.3 | 46.9 |
| AM10 | 5.04ab | 8.8 | 5.6 | 30.1 | 1.7 | 43.2 | 1.5 | 22.2 | 45.7 |
| AM11 | 5.01b | 15.5 | 5.3 | 29.1 | 1.5 | 47.0 | 2.0 | 23.7 | 43.3 |
| AM12 | 5.06ab | 15.5 | 5.4 | 27.7 | 1.5 | 47.4 | 2.3 | 26.4 | 43.3 |
| AM13 | 4.94b | 18.4 | 7.9 | 27.1 | 1.6 | 47.9 | 2.2 | 25.9 | 43.7 |
| MB– Microbial Biota | |||||||||
| MB1 | 5.08a | 12.1 | 7.4 | 28.4 | 1.4 | 42.6 | 1.9 | 20.2 | 46.3 |
| MB2 | 5.12a | 14.7 | 8.4 | 28.3 | 1.4 | 44.1 | 1.9 | 21.1 | 46.7 |
| C– Control | |||||||||
| C | 5.18a | 6.7 | 5.6 | 25.0 | 1.3 | 45.5 | 2.3 | 25.8 | 43.9 |
| MB/C | -1% | 99% | 42% | 13% | 10% | -5% | -18% | -20% | 6% |
| Prob | 0.03 | - | 0.01 | 0.00 | 0.37 | 0.24 | 0.01 | 0.00 | 0.019 |
| AM/C% | -3% | 129% | 20% | 12% | 18% | 2% | -8% | -6% | 1% |
| Prob | <.0001 | - | 0.20 | <.00 | 0.00 | 0.94 | 0.13 | 0.34 | 0.80 |
| Thesis | Ash % | In vitro Digestibility % | NDF non dig. % | NDF digestibility % | ADL % | NDF % | Crude Protein % | Cellulose % | Dry Matter %£ |
| AM – Arbuscular Mycorrhizae | |||||||||
| AM01 | 8.0 | 70.4 | 29.6 | 42.3 | 6.3 | 54.5 | 6.7 | 22.1 | 32.6 |
| AM14 | 8.7 | 72.0 | 28.0 | 47.3 | 8.8 | 50.9 | 9.1 | 19.9 | 30.3 |
| AM06 | 7.6 | 71.1 | 28.9 | 45.6 | 8.8 | 49.6 | 8.7 | 18.2 | 28.9 |
| AM02 | 10.1 | 72.5 | 27.5 | 52.3 | 8.4 | 51.1 | 10.1 | 17.5 | 29.7 |
| AM09 | 9.5 | 71.6 | 28.4 | 44.6 | 8.4 | 51.4 | 9.3 | 19.1 | 28.9 |
| AM03 | 9.1 | 71.8 | 28.2 | 43.1 | 7.9 | 51.2 | 9.6 | 22.4 | 29.4 |
| AM12 | 9.6 | 71.4 | 28.6 | 44.1 | 8.5 | 51.6 | 9.3 | 18.8 | 29.7 |
| AM10 | 10.3 | 72.2 | 27.8 | 44.9 | 7.8 | 52.2 | 11.5 | 24.5 | 29.3 |
| AM05 | 7.3 | 72.0 | 28.0 | 46.1 | 8.8 | 54.1 | 9.8 | 22.4 | 29.0 |
| AM11 | 10.8 | 76.6 | 23.4 | 52.9 | 8.2 | 50.4 | 9.5 | 19.7 | 28.8 |
| AM08 | 8.5 | 71.1 | 28.9 | 40.3 | 7.0 | 52.4 | 8.3 | 20.6 | 31.0 |
| AM13 | 7.7 | 69.6 | 30.4 | 43.3 | 9.5 | 52.1 | 8.3 | 21.0 | 33.0 |
| AM07 | 8.6 | 70.9 | 29.1 | 43.7 | 8.0 | 53.3 | 9.6 | 22.2 | 30.2 |
| MB - Microbial Biota | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| MB1 | 10.5 | 72.9 | 27.1 | 49.4 | 7.3 | 49.3 | 10.5 | 23.8 | 29.4 |
| MB2 | 10.3 | 72.7 | 27.3 | 47.7 | 6.8 | 53.0 | 10.4 | 22.9 | 30.8 |
| C – Control non-inoculated | |||||||||
| C | 7.1 | 70.0 | 30.0 | 42.3 | 7.5 | 52.7 | 7.1 | 23.6 | 33.6 |
| MB/C% | 46% | 4% | -10% | 15% | -7% | -3% | 48% | -1% | -10% |
| Prob | 0.00 | 0.06 | 0.07 | 0.13 | 0.37 | 0.57 | 0.01 | 0.99 | - |
| AM/C% | 25% | 3% | -6% | 7% | 9% | -1% | 30% | -12% | -11% |
| Prob | 0.03 | 0.12 | 0.12 | 0.48 | 0.78 | 0.89 | 0.03 | 0.03 | - |
Figure 1. Histogram of the % decrease in the raw pH of the inoculated theses from the non-inoculated Control, ordered by values.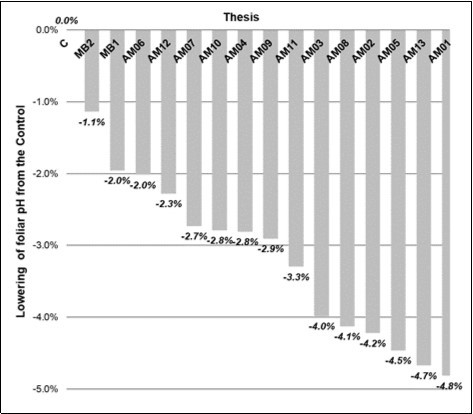
As far as the biofertilizers are concerned, MB1 (-1.1%) was more acidifying than MB2 (-2.0%), and the AM were scaled according to three gradients of acidity: the most acidic strains, that is, below -4%, were the AM01, AM13, AM05, AM02 and AM08 strains.
NIR-Tomoscopy, Foliar Weight and Composition
The weight of the leaves harvested from the plants was much more reduced in the non-inoculated Control than in all other theses (Table 3a). Moreover, the foliar composition was affected by the category of the inoculant, as calculated from Table 3a and Table 3b. The MB category scored 7 significant increments and 3 decrements vs. the C, while the AM category scored 5 and 1 variations, respectively.
As can be observed in Figure 2, the constituents augmented in the leaves of the inoculated Sorghumsudanensis were: crude protein, ash, digestible NDF, hemicellulose and the ether extract; the constituents reduced in contents and properties, namely dry matter, crude fiber, cellulose and crop maturity index, were increased less.
Figure 2. Histogram of the % deviation in the foliar composition of the AM and MB categories from the non-inoculated control, ordered according to the AM values.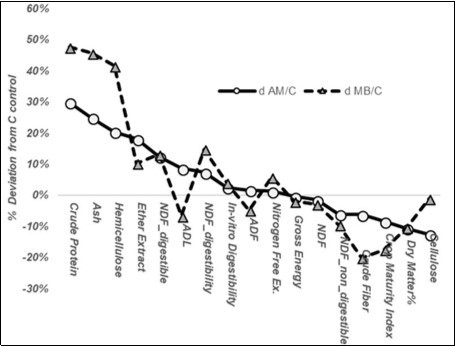
Foliar pH as the Key-Variable
The PLS standard model (Figure 3), with R2=0.81, showed that a low pH steadily favoured the dry mass weight and, to a lesser extent, the hemicellulose, digestible NDF and ether extract contents; on the other hand, a high pH tendentially increased the dry matter percentage and the cellulose content of the leaves.
Figure 3. Histogram of the normalized Partial Least Squares regression coefficient of the raw foliar PH
NIR-Tomoscopy
In general, the discriminations of the three categories when the SCIO instrument was used were highly significant (Table 4). The leaves from plants inoculated with AM were individuated more correctly, in the scans on the fresh leaves (96%) as well as in the scans on the dry leaves (94%); the performances pertaining to the MB leaves were significant, albeit not so high (59 and 51%, respectively), while the Control leaves were individuated more clearly (74 and 65%, respectively).
Table 4. Reclassification of the AKA matrices of the NIR-SCIO spectra from fresh and dry leaves into three Categories, and Probability of the diagonal elements.| NIR- Tomoscopy (No. 480) | Dry leaves (No. 48) | ||||||
| Categories | C | MB | AM | C | MB | AM | |
| Arbuscular Mycorrhizae (AM) | 22% | 31% | 96% | 30% | 48% | 94% | |
| Microbial Biota (MB) | 5% | 59% | 2% | 5% | 51% | 3% | |
| Control (C) | 74% | 11% | 1% | 65% | 0% | 1% | |
| Prob. Diagonal elements | P | < 0.0001 | < 0.0001 | < 0.0001 | < 0.0001 | < 0.0001 | < 0.0001 |
Discussion
Mycorrhizal Modifier
A luxuriating effect has been displayed in foliar development. A meta-analysis of the field studies on the responses of wheat to AM has highlighted that field AM inoculation can be proposed as an effective agronomic practice for wheat production7, with aboveground biomass increases of around 20%, as already assessed in maize under high11,15, 22, 23, 24or low25Italian input conditions and under Indian conditions21
As expected, a more juvenile ontogenic status emerged for the leaves of inoculated plants but graded among the inocula types.
Claps et al.26, in field conditions, after the autumn sowing of barley (Hordeumvulgare) and clover (Trifoliumalexandrinum) tanned at 1 kg ha-1 with the MB1 of the present work, observed a slight increase in yield (not significant), but also significant variations in composition (Figure 4), albeit only in the barley grass. As in the present work, the advantage was centred on crude protein (+18%), but in that case a strong reduction in fibrousness was found, which enhanced the digestibility coefficients of the NDF, crude protein and organic matter. On the other hand, a reduction in ash (-10%) disagreed with the increase observed in the present work.
Figure 4. Histogram of the % deviation in the grass composition of the treated barley, according to Claps et al.26, with MB1 from the non-inoculated control, ordered according to the values.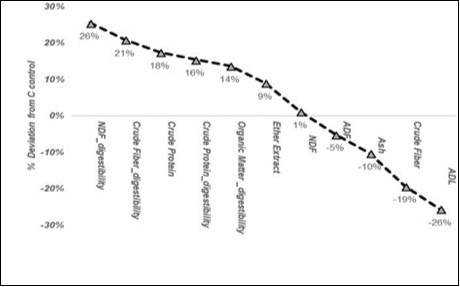
Other AA have conducted experiments on AM at more mature stages, but hay and silages have rarely been studied. Uzun22 observed that no reductions in forage yield occurred in Sorghumsudanensis or maize, when the diminished supply of fertilizers was balanced with the action of Glomus spp. and the forage produced using an AM treatment showed better chemical characteristics when harvested at an appropriate phenological stage.
As far as maize forage is concerned, Sibi et al.27 observed that an AM fungus was significant, at a level of 5%, on water soluble carbohydrates, which increased by 14%. The Authors hypothesised that inoculation with an AM fungus increased the cytokinins, and, moving nutrients from other parts to the leaves tended to delay their senescence.
Raw pH fall-out as a Key-Variable
In our short modification experiment, low pH steadily favoured the dry mass weight and, to a lesser extent, the hemicellulose, digestible NDF and the ether extract contents of the leaves. If the raw pH is a real key-variable, the modifier direction in Sorghumsudanensis appears quite attractive for ruminants. Phyto-evolution studies17,28 of a subarctic flora suggest that tissue pH itself is controlled to a great extent for a given species, because of its direct or indirect functions in the plant; in fact, a low pH corresponds to poor digestibility and may therefore act as an anti-herbivore defence in the same subarctic flora.
The field surveys of AM communities over a wide range of soil pH suggest that it is also the major driving force in structuring these communities29.
It was found, in the microcosm experiments6, that N2O emissions were cumulatively 42.4% higher without AM than with AM microcosms. However, it should be recalled that the Authors, in their tomato experiment, observed an increase in soil pH after an AM inoculum of 7.75 vs. 7.62 (+1.6%; P=0.004). Unfortunately, the raw pH was not measured in that experiment. In light of our present and previous results15,16, it is possible to hypothesize that the transfer of [H+] from the soil to the AM plants could have effectively reduced the circulating acidity of the soil. Several results support this hypothesis. Bago et al.30 demonstrated, in a sterile culture with Glomus intraradices, that extra-radicalmycelium utilised NO3-N and increased pH of the medium by up to one unit. Li and Christie31 showed that soil solution Zn concentrations were lower and pH values were higher in mycorrhizal treatments than non-mycorrhizal controls, and the AM effects were more pronounced at higher Zn application rates.
Vibrational Spectroscopy
The direct NIRS discrimination of bio-fertilized crops is rare. A first work on Ocimumbasilicumcv. “Genovese”32 assessed that three AMs were modifiers of growth, and distributors of glandular trichomes and of the essential oil composition, with a homologous relative pattern that was appreciated by NIRS and Electronic Nose. A smart NIRS technique that has recently been applied to hay-litter-bags19has produced promising results in discriminating the fingerprint effects of bio-fertilizers. The results of direct NIR discrimination have instead been obtained for several cases. Szuvandzsiev et al.33 suggested that the effect of very different irrigation regimes on tomato leaves is detectable by means of NIRS, irrespective of the cultivar and phenological stage; around 960 nm emerged a good correlation between different irrigation regimes and the reflectance of the raw spectra of tomato leaves taken from the 900-1000 nm representative spectral range. Foliar UV-Vis-NIR spectra (325-1075 nm) was used to discriminate high oil and high protein from a normal maize type on five sampling dates over two years34; the results showed that the qualitative models constituted a robust classification method; however, the best method on average attained only 58.3±5.5% of validation, a superior value to the hazard threshold of 33%, but of no use for traceability purposes. The somewhat optimistic values reported in the present work, obtained using the smart NIR-SCIO (Tab. 4), need to be confirmed over some years and under different conditions.
Conclusions
In a meta-analysis concerning nearly half a million species x sites worldwide on twenty-one plant traits, Moles et al.35stated that the mean annual temperature was correlated more with the plant traits than with the mean annual precipitation. Considering the present results, a further recommended easy trait that could be considered is the raw pH.
In this work, the fall out of the raw pH responded as a simple sign of AM inoculation and activities. However, various responses were observed for the same Sorghum sudanensis with different AMs, thus suggesting more useful combinations and interactions, where the raw pH can be considered a key-variable of plant modifications in an AM framework.
The vibrational spectroscopy of leaves can be used to rapidly classify the outcomes linked to symbiotic agents, and a smart network can be used to capitalize on useful information in an NIRS leaf data-base.
From a practical point of view, it may be recommended to monitor raw pH for traceability purposes in an AM multiplication framework.
Pursuing the study of pH from a plant-to-soil point of view is better than the inverse, and each search conducted to connect the microbial soil fingerprint (hay-litter-bags19or others36)to the NIRS fingerprint and productivity traits of leaves can be considered useful.
Acknowledgments
The authors wish to thank the Fondazione CRT, Torino – Italy, for the financial support to the scientific activities of the “Accademia di Agricoltura di Torino”.