Abstract
Effects of the plant lectin phytohemagglutinin M (PHA-M) and illumination on the electrophoretic mobility (EPM) and 90°-light scattering of suspension of spheroplasts obtained by lysis of the unicellular diazothrophic cyanobacteriumPlectonemaboryanumwere studied. Cells cultures were grown on media with different iron content: an iron-deficient (denominated as “Fe-starved” culture), an iron-sufficient (“Fe-sufficient”), and a twentyfold iron excess medium (“Fe-excess”). Lectin addition led to a decrease of the membrane charge density and aggregation of the vesicles. Our results indicate significant influence of the Fe2+ and illumination on the lectin-induced decreasing of EPM and light scattering intensity.
Author Contributions
Academic Editor: Qian Zhu, School of Informatics and Computing , Indiana University Bloomington. USA
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2014 Virjinia Doltchinkova, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Competing interests
The authors have declared that no competing interests exist.
Citation:
Introduction
The electric charge distributed on the surface of biological membranes plays an important role in the regulation of the molecular membrane processes, but also in the processes of interaction of biological cells with chemical substances. Deprivation of micronutrients can profoundly affect photosynthetic electron transfer 1. Iron (Fe), manganese (Mn), magnesium (Mg) and copper (Cu) are essential cofactors for the operation of the oxygenic photosynthetic electron transfer apparatus2. Although abundant in the earth’s crust, Fe predominates in insoluble Fe (III) precipitates and is largely unavailable to plants, especially at neutral or alkaline pH.
As most biological membranes, spheroplasts isolated in vitro as closed vesicles are negatively charged at neutral pH. Investigation of the nature of the surface charges is important in order to obtain information about the surface components and the aggregation processes of the spheroplasts vesicles. Using the Gouy-Chapman theory3, 4,we investigated the influence of chemical modification of negative charges on the thylakoid membranes and found that they facilitate the entropy-driven ‘depletion attraction’ between the adjacent membranes.
The widespread distribution of lectins throughout the plant kingdom and their abundance in many plants suggest that these molecules are of physiological importance5. Lectin receptors are the kind of protein macromolecules playing a role of specific determinants of cell-cell recognition. The characteristic property of lectins is their ability of binding to specific carbohydrate structure and to provide by that a basis of biologically reliable recognition6. Due to their multivalent binding character the lectins, when added exogenously, cause crosslink and molecular reorganizations in the cell membrane bilayer7.
Phytohemagglutinin (lectin of Phaseolus vulgaris agglutinin) activation of aggregate processes serves as a model to study the biophysical mechanisms of cells aggregation8 .Light-induced processes of aggregate formation of spheroplasts included in complexes were registered after illumination with intensity of 920 μmol quanta m-2 s-1.
In this work we study electrophoretically the galacto residues on the outer membrane surface (enabling the interaction with other plasma membranes) in dependence of iron content in the growing culture. Respectively, the spheroplast suspensions are designated as: ‘Fe-starved’ (absence of Fe2+ in the medium), ‘Fe-sufficient’ (37.2 μg/L Fe2+) and ‘Fe-excess’ (744.9 μg/L Fe2+, twentyfold increased). The electrophoretic mobility (EPM or u) of Pl. boryanumspheroplasts were measured by microscopic (visual) microelectrophoresis. The light scattering method was employed for determination of the aggregate mass in “Fe-sufficient” and “Fe-excess” suspensions of spheroplasts upon light and lectin treatment; the significance of these techniques at strong aggregation is discussed.
The combined employment of electrokinetic and light scattering methods allows both analyzing the surface components of the plasma membrane and the spheroplast aggregate formation processes. In this study we report for the first time: 1) light-induced increase in EPM, ζ potential and surface electric charge of ‘Fe-sufficient’ spheroplasts 2) increase in aggregate formations upon light of lectin pre-treated spheroplasts in dependence on iron content in growing media of Pl.boryanum.
Materials and Methods
Materials
Chemicals were analytical grade. Bacto Phytohemagglutinin M (PHA-M), M.w. 124 kDa, was purchased from DIFCO Laboratories (Detroit, Michigan, USA). Hepes (N-(2-Hydroxiethyl) piperazine-N-(2- ethanesulfonic acid) and NaCl were purchased from Merck. K2HPO4 and KH2PO4 and BSA (Albumine bovine, fraction V) were obtained from Sigma. Lysozyme from chicken egg white 20.000 units/mg was obtained from Serva Feinbiochemica (Heidelberg/New York).
Cultivation of Plectonema Boryanum
Pl. boryanumGom. (Leptolyngbya boryana (Gomont), Anagnastidis et Komarek, strain 594, is a filamentous nonheterocystous diazotrophic cyanobacterium that was grown autotrophycally on the nutrition medium9. The culture was cultivated in iron-free culture (“Fe-starved”) and at two different iron concentrations of Fe as the EDTA complex in the nutrition medium by using metal salt FeSO4.7H2O (Fe2+): 4.00 ppm metal (“Fe-sufficient”), and 20 times higher quantity of iron (80.00 ppm metal, “Fe-excess”). After 48-hour cultivation the algal cells were harvested at the end of the exponential phase of growth by filtration and washed twice with distilled water.
Preparation of Spheroplasts
The culture of P. boryanum at a logarithmic phase was centrifuged for 20 min at 5 000 x g. The sedimented cells were suspended in 30 mM K2HPO4/KH2PO4 buffer (pH 7.0) and the cells were collected by centrifugation as above. For preparation of spheroplasts, the cells were suspended in 50 mL of 1 mg/mL lysozyme, 0.5 M sucrose, 2% BSA, 30 mM K2HPO4/KH2PO4 buffer (pH 7.0) and then incubated in dark for 4 hours at 37 °C on water bath shaker ‘Elpan’, type 357. The lysozyme treatment was stopped by centrifugation for 10 min at 2 240 g at 4 oC. The spheroplasts were washed and sedimented twofold by centrifugation for 5 min at 2 240 g at 4 oC in a buffer, containing 0.5 M sucrose, 2% BSA and 30 mM K2HPO4/KH2PO4 buffer, pH 7.0. Immediately after stirring of sedimented cells in 1 mL of the upper buffer, the spheroplast suspension was cooled at 0 oC in dark10, 22. The final intracellular concentration of chlorophyll a determined spectrophotometrically after methanol extraction11 was about 6 μg/ mL.
The spheroplast formed in this way had a microscopically estimated size of about 3 μm and mean volume of 500 μL per vesicles. The spheroplasts suspensions were incubated for 30 min at 23 °C without or in the presence of PHA-M (10-7-fold diluted in an 25 mM Hepes buffer, pH 7.5, 10 mM NaCl), with final concentration 2-40 pg/mL. The preparation was stored on ice in dark until use (within 4 h).
For light scattering (LS) and electrophoretic measurements were used samples with 10 mM NaCl, 25 mM Hepes (KOH) buffer, pH 7.5. The electrical conductance and viscosity of the different suspensions were measured using a CyberScan PC 510 (Eutech Instruments, USA, and Singapore) pH/Conductivity meter and a Rheo (Germany) viscometer, respectively.
Microelectrophoresis
The samples for electrophoretic measurements were prepared by mixing of 200 μL spheroplasts suspension (with appropriate concentration of lectin) and 20 mL of buffer (consisted of 25 mM Hepes (KOH), 10 mM NaCl, pH 7.5).The stock solution of PHA-M was prepared in doubled distilled water.
The electrophoretic mobility (EPM) measurements were performed using the particle electrophoresis technique OPTON cytopherometer (Feintechnik Ges, m.b.H., Wien, Austria). Electrophoretic migration was measured in a rectangular chamber with platinum electrodes at a constant d.c. electric field with strength 100 V/cm, the current in the cell was 3 mA. The measurements were implemented at 25 oC. The movement of 15-25 spheroplasts moving over a distance of 32 μm was timed using microprocessor equipment for both forward and backward (reversed field) runs. The observation light (with intensity of 13 μmol quanta m-2 s-1) was filtered through a green (V G 9 (46 78 05) 545 nm interference filter which improves the contrast of the picture details. The spheroplasts suspensions (chlorophyll a concentration of 6μg mL-1) were illuminated for 1 minute at 25°C in the electrophoresis chamber with actinic light intensity of 920 μmol quanta m-2 s-1 by the lower voltage lamp (Wotan 38 01 77 Germany 6 V 15 W iYG). The spheroplasts were observed with Sony video camera providing 800 times magnification. The data were averaged over 65-80 measurements for three independent samples at every lectin-Fe concentration and presented as (u ± standard deviation) in units [10-8 m2V-1s-1].
The zeta potential (ζ) was calculated from the electrophoretic mobility, u, using Helmholtz-Smoluchowski equation12 :
where eris the relative dielectric permittivity of the aqueous phase (er = 78.5 at 25 °C), e0 is the electric constant in SI (e0 = 8.85 x 10-12 F m-1), η is the viscosity of the liquid phase (η = 1.21 ± 0.02 mPa×s for solution of 25 mM Hepes buffer and 10 mM NaCl).
When the spheroplasts are suspended in a medium containing a symmetric electrolyte (Z:Z, such as NaCl or MgSO4) with valency z and concentration Ci the surface charge density s is:
ϭ = 0.1174 Ci 1/2 sinη (zψ/51.7)(2)
where ϭ is in C×m-2, Ci is in mol×L-1 and ψ is in mV; ψ ≈ζ 13.
Statistical analysis: Significant differences between means were determined by use of Analysis of Variance (ANOVA). Kruskal-Wallis One-Way Analysis of Variance on Ranks was performed with Holm-Sidak method for comparison the significance of the treatment as well as Dunn’s method for all pair wise multiple comparison procedures taking p≤0.05 as significant.
Light Scattering Measurements
The aggregation of the spheroplast suspension was studied by intensity of the light scattered at 90° and wavelength 480 nm. For this purpose Specol 10 Spectrophotometer (Carl Zeiss, Jena) with fluorescent attachment was used. The light scattering intensity (LS) is measure for the degree of aggregation of the spheroplasts.
The samples were prepared by addition of spheroplasts suspension (6 mg chlorophyll per mL) to 20 ml reaction medium and following incubation at 25°C for 3 min.
LS of the probe was measured after illumination with white light by 6V,15W lamp. The LS response of the spheroplast membranes consisted of a main phase with signal amplitudes in dependence of lectin pre-treatment applied on the spheroplasts
Statistical analysis: Data were averaged of triplicate measurements for every sample.
The significant differences between means were determined by use of ANOVA. One-Way Analysis of Variance was performed with Holm-Sidak method for comparison the significance of the treatment as well as Student-Newman-Keuls method for all pair wise comparisons on the mean responses among the different groups taking p≤0.05 as significant.
Results
Phytohemagglutinin–Induced Changes of EPM of Spheroplasts
The technique of particle microelectrophoresis was used to measure the electrophoretic mobility u of spheroplast suspended in low ionic strength media (0.01 M). The surface charge density was estimated from u.13
In the first set of experiments spheroplasts suspension from all the variants was exposed to a white light treatment for 1, 2, 5 and 10 minutes. There was a significant change both in the electrophoretic mobility and the basal light scattering evaluated with respect to the Control point without illumination (0 min point). The light treatment duration was selected to be 1 min. The electrophoretic mobility of “Fe-sufficient” spheroplasts significantly increased after 1 min light exposure which was accompanied by a reduction of more than 12% in LS of the same particles in a buffer of low ionic strength at temperature (25 oC). Illumination (2 min, 5 min and 10 min) led to a significant decrease in EPM by 18.5% (p=0.002) and 16% (p<0.001), respectively. Light treatment (2 min and 10 min) of “Fe-starved” spheroplasts induced a strong enhancement of EPM up to 43.3% and 20.7%, respectively (p<0.001).There were no significant changes in “20 x Fe” spheroplasts after light exposure for all the minutes of treatment. There were structured aggregates of compactness spheroplasts with reorientation during their electrophoretic mobility in d.c. electric field. The “20 x Fe” spheroplasts possess a high polarity and separation and reorientation of some particles in the aggregate during movement in electric field were observed. Some of these particles became more “heavy” upon 10 minutes illumination without a significant change in light scattering of the “20 x Fe” spheroplasts. They changed the direction of movement in electrophoresis chamber. The EPM of these particles were not included in the EPM data of the upper variant.
We studied the electrokinetic and light-scattering effect in spheroplast membranes up to 4 hours after storing the spheroplast suspension on ice.
Lectin had no significant effect on EPM of the “Fe-sufficient” spheroplast particles; only in the case of treatment by dose of 40 pg/mL PHA-M the mobility decreases with 16% Figure 1 A, p<0.001). But the EPM of “Fe-starved” spheroplasts is some higher at middle lectin concentrations; the increasing reaches 23% at 10 pg/mL PHA-M. SDS-PAGE showed new electrophoretic bands of 55.5 kDa and 57.1 kDa proteins were appeared at “Fe-starved” Pl. boryanumvariant (data not shown). The data obtained indicated that accumulation of these complexes could be regarded as a manifestation of compensatory mechanism activity under conditions of iron starvation.
After illumination EPM of the “Fe-sufficient” spheroplasts increases at high lectin concentrations; the increase reaches 43% at 40 pg/mL PHA-M (compared to EPM of illuminated ones without lectin addition) Figure 1 B. As shown in Table 1, the surface of “Fe-sufficient” spheroplasts became more negative as observed by an increase in EPM at 10 - 20 pg/mL PHA-M and 40 pg/mL PHA-M compared with lectin pre-treated spheroplasts before light exposure. In the case of “Fe-starved” the illumination has more discrepant effect: EPM decreases at middle lectin concentrations and increases at high ones (up to 27% at 30 pg/ mL PHA-M).
Figure 1. Relative electrophoretic mobility (EPM) of (A) spheroplasts treated by PHA-M or (B) pretreated by PHA-M and then illuminated. The medium contained 25 mM HEPES (KOH) buffer and 10 mM NaCl at pH 7.5. Data are means of three independent experiments (n=50-70 measured spheroplasts). EPM values measured in the absence of PHA-M are equated to 100%. The spheroplasts are prepared from the culture of Plectonema boryanum grown at absence (Fe-starved culture, light red columns) or in the presence of iron in the cultivating medium at concentrations: 37.2 μg L-1 Fe2+ (Fe-sufficient culture, white columns) and 744.9 μg L-1 Fe2+ (Fe-excess culture, black columns).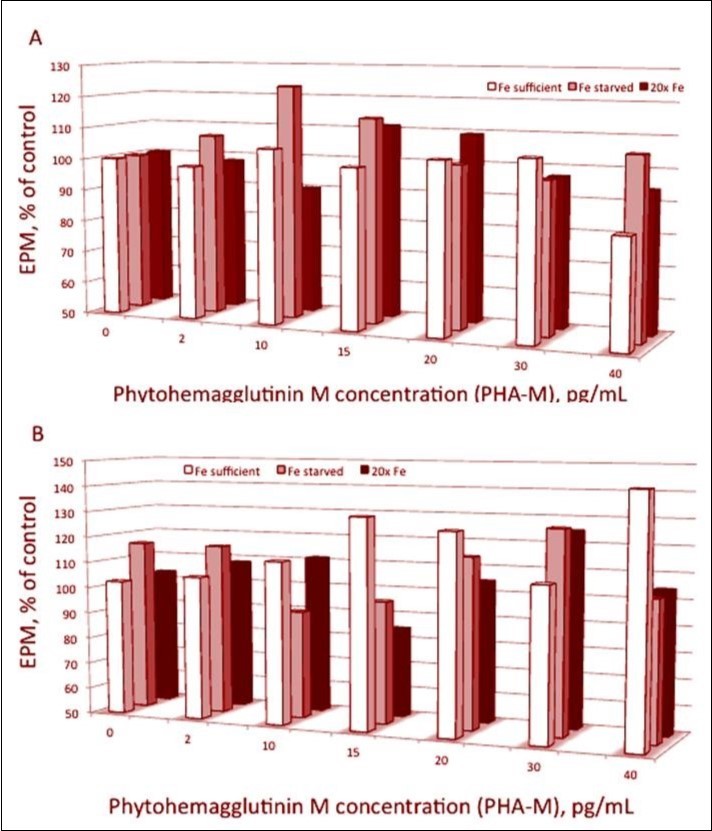
| Electrophoretic mobility u x 108 (m2V-1 s-1± SEM)before illumination | |||||||
|---|---|---|---|---|---|---|---|
| Lectin Variants | Control0 pg PHA-M/mL | 2pg PHA-M/mL | 10pg PHA-M/mL | 15pg PHA-M/mL | 20pg PHA-M/mL | 30pg PHA-M/mL | 40pg PHA-M/mL |
| Fe-sufficient (n=17-24) | -1,55±0,05 | -1,52±0,05 | -1,56±0,04 | -1,55±0,05 | -1,60±0,05 | -1,62±0,07 | -1,30±0,07***p˂0,001 |
| Fe-starved (n=15-19) | -2,01±0,05 | -2,18±0,06**P=0,037 | -2,56±0,03***p˂0,001 | -2,33±0,06***p˂0,001 | -2,13±0,08 | -2,04±0,08 | -2,15±0,06 |
| 20 x Fe (n=15-21) | -0,99±0,05 | -0,97±0,05 | -0,89±0,04***p˂0,001 | -1,10±0,07 | -1,08±0,06 | -0,96±0,06 | -0,93±0,04 |
| after illumination | |||||||
| Fe-sufficient (n=18-23) | -1,58±0,03 | -1,60±0,06 | -1,81±0,05**p=0,026 | -2,02±0,05***p˂0,001 | -2,02±0,06***p=0,003 | -1,76±0,05 | -1,85±0,05**p=0,008 |
| Fe-starved (n=16-21) | -2,33±0,06 | -2,36±0,03 | -2,30±0,06 | -2,26±0,05 | -2,25±0,04 | -2,52±0,04 | -2,39±0,06 |
| 20 x Fe (n=15-20) | -1,02±0,06 | -1,05±0,04 | -0,99±0,06 | -0,93±0,13***p˂0,001 | -1,13±0,04 | -1,20±0,04**p=0,028 | -0,97±0,018***p=0,005 |
Lectin-Induced Aggregation of Spheroplasts
In the second set of experiments the effect of light exposure on the lectin-induced aggregation behavior of spheroplast vesicles were determined as a function of PHA-M concentration. The light exposure did not change visually the structure of the aggregates of spheroplast after lectin treatment.
LS was decreased upon rising lectin concentration in all the variants of non-illuminated spheroplasts Figure 2, p<0.001. Strikingly, at the decrease appears at low concentration of PHA-M in the case of “Fe-sufficient” spheroplasts as opposed to high PHA-M concentrations when the cultures were grown in lack or in 20-fold excess of Fe2+. The aggregation behavior of illuminated spheroplasts is different: LS does not depend on the lectin concentration at lack of sufficient amount of soluble iron in the growing medium. Therefore, further investigations could be carried out with an aim to: (1) aggregation of spheroplasts under iron stress and (2) investigate the nature of the LS changes in dependence of iron content in suspending media of Plectonema boryanum.
Figure 2. Light scattering of spheroplast vesicles as determined by basal LS in the absence (=100%) and in the presence of various concentrations of PHA-M. Spheroplasts and various quantities of PHA-M were suspended in a medium containing 25 mM HEPES (KOH), 10 mM NaCl, pH 7.5. Chlorophyll concentration was 6 μg mL-1. Relative LS of PHA pre-treated spheroplasts before light expose. Data are means of three independent measurements (9 measurements at given concentration of iron and PHA-M).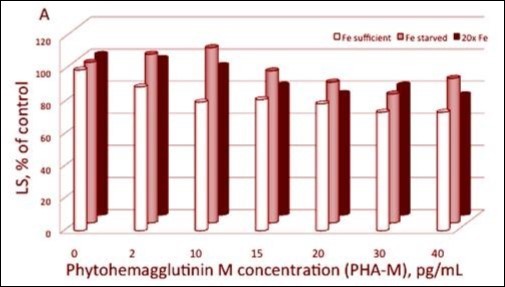
Discussion
The experiments presented in this study aim to characterize the electrical properties of photosynthetic membranes isolated from Pl. boryanumcyanobacterium when the culture is grown at lack, sufficient or excess of Fe2+ in the culture medium. For this purpose the methods of microelectrophoresis and light scattering are used giving information about surface charge density and aggregate’s mass, respectively. Changes of the electrokinetic mobility and aggregation behavior of the spheroplasts are caused by addition of lectin to the suspension and by light illumination causing photosynthetic reactions in their membrane. The biological effect of light has been widely studied. We used the microscopic (visual) microelectrophoresis as the more convenient method for determining the electrophoretic mobility of the spheroplasts.We applied the Gouy-Chapman theory that has been developed to describe the ions distribution in the diffuse electrical layers associated with charged surfaces14. As the spheroplast membrane is negatively charged at neutral pH, cations tend to be electrostatically drawn close to the membrane for the sake of neutrality and form a diffuse layer.
In this study the effect of polyvalent ions was tested by measuring the EPM at pH 7.5 in the presence of different concentrations of lectin phytohemagglutinin M (PHA-M) with molecular mass of 124 kDa. PHA-M as a tetrameric protein with 4 sugar-binding sites could manifest an ‘effective’ positive charge of 2.2 at 0.01 M ionic strength15. The binding of PHA-M macromolecules to the N-acetyl-D-galactosamin oligosaccharide residues of the membrane glycolipids was reported16 . These properties of PHA-M determinate a specific binding of the lectin macromolecules to membrane what reduces its surface charge density. Because of the strong affinity of a given lectin to specific polysaccharides of the glycolipids its binding appear at very low concentration but after saturation of the binding sites on the membrane surface further increasing of the lectin concentration leads to electrostatic (non-specific) adsorption at much higher concentration as in the case of simple ions not having anti-body properties. One example is the binding of PHA to the negatively charged purple membranes17 ; that cause decreasing of the EPM and alteration of their curvature because of asymmetric binding 18, 19.
Our results showed that there are no significant differences in the EPM of non-illuminated spheroplasts at lectin concentrations up to 30 pg/mL PHA-M in the case of Fe2+-sufficient culture (Figure 1 A). That means that at normal conditions the cyanobacterium cells do not synthesize the polysaccharide antigens on the surface of their photosynthetic membranes.
Our results shows out that the lectin binding involved vary from variant to variant in a specific manner; that gives impression of irreproducibility of the results. Figure 1 suggests that EPM as a function of PHA-M concentration have some periodicity but different at the three kind of spheroplasts (obtain from cultures with different content of Fe2+). How the EPM depends on the iron concentration in the culture medium, remains to be determined but the electrophoretic behavior of the spheroplasts remember the electric properties of chloroplasts and subchloroplast vesicles obtained from higher plants as peas20, 21 . Upon treatment with dose of the 40 pg/mL PHA-M a reduction in the net negative surface charge density of “Fe-sufficient” spheroplasts (from – 0.0046 C m-2 without lectin to – 0.0038 Cm -2) was observed. A significant increase in σ of “Fe-starved” spheroplasts (from – 0.0060 C m -2 without lectin to – 0.0078 C m -2 post 10 pg/mL PHA-M) was derived.
The electrostatic effect of PHA-M was slightly expressed due to unspecific binding of lectin to outer surface of the plasma membranes of Pl. boryanumspheroplasts. Only 10 pg/ mL PHA-M significantly increased the net negative surface charge density because of structural modification and rearrangement.
The influence of light exposure on the surface charge density s was manifested by the changes in binding of lectin (at PHA-M concentrations where electrostatic interactions predominate) to plasma membranes of spheroplasts. An increase in s of “Fe-sufficient” spheroplasts after illumination from – 0.0047 C m -2 without lectin to – 0.0060 C m -2 in the presence of 15-20 pg/ mL PHA-M was observed. There was a maximal increase in surface electrical charge after 40 pg/mL PHA-M treatments (Figure 1, B). The effect of illumination is better expressed at the normal content of Fe2+ in the growing culture; that suggests influence of the iron on the biochemical processes of formation of the photosynthetic complex in the membrane.
The influence of Fe2+ on the membrane electric properties was also studied by the aggregation behavior of the spheroplasts in suspension. Figure 2 shows that LS intensity decreases upon PHA-M treatment but differently for the three types of culture. The lectin’s effect appears at significantly lower concentration at “Fe-sufficient” spheroplasts than in the cases of lack and twentyfold excess of Fe2+ in the culture medium. Those results confirm the above made suggestion about the importance of the iron ions for proper formation and functioning of the photosynthetic membranes.
Acknowledgements
This work was partially supported by National Science Fund, Ministry of Education, Youth and Science, Bulgaria with grant number (HS-L-301/07). We would like to thank Assist. Prof. Ganka Chaneva (Department of Plant Physiology, Sofia University “St. Kliment Ohridski”) for generously providing the Pl. boryanumstrain and to Assist. Prof. Teodora Georgieva (Department of Genetics, Sofia University “St. Kliment Ohridski” for SDS PAGE analysis.
Abbreviations:
chlorophyll
LSlight scattering
EPMu
ζzeta (electrokinetic) potential
σsurface charge density