Abstract
Antibody phage display has become a useful technique for discovering and optimizing target-specific monoclonal antibodies suitable for many applications, including therapeutic ligands, which may act as direct pharmacological compounds or may be used as targeting ligands for controlled drug delivery. Recently, the D2-5-HT1A heteromer, which is formed by the dopamine D2 and serotonin 5-HT1A receptors has attracted attention as a potential target of antipsychotic drugs. Therefore, the aim of the study was to identify scFv monoclonal antibodies that are able to specifically recognize epitopes formed within the heteromer structure. Because both receptors are membrane proteins, it is important to conduct bio-panning experiments in the most natural conditions, in which the presented antigens (D2-5-HT1A heteromers) are in their native form and possibly in their best-preserved spatial structure. It has been shown here that phage display methodology can be successfully used in the preparation of monoclonal antibodies against dimers of membrane proteins. To separate phages specifically binding the D2-5-HT1A heteromer, the selection process using CHO+ cells with overexpression of both receptors was conducted. Phages that were bound to receptor monomers or other CHO-K1 cell surface proteins were eliminated as a result of negative selection by using CHO- cells expressing separate receptor monomers.
Author Contributions
Academic Editor: Xufen Yu, Icahn School of Medicine at Mount Sinai, United States.
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2019 Sylwia Łukasiewicz, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Competing interests
The authors report no conflicts of interest in this work. The authors also declare no competing financial interest.
Citation:
Introduction
Nanoparticulate systems with functionalized surfaces for targeted drug delivery play central roles in modern therapies 1, 2, 3. In our previous studies, we focused on the encapsulation of clozapine (one of the most important antipsychotic drugs) into biocompatible polymeric nanocapsules formed by polyelectrolyte multilayer shells 4, 5. Promising nanocarriers able to cross the blood brain barrier (BBB) were obtained 6; however, to achieve higher selectivity, additional functionalization of the nanocarrier is needed. This necessity is well exemplified by clozapine, which is effectively used in the clinic; however, clozapine displays many serious side effects that most likely could be eliminated if we knew the precise mechanism responsible for its efficacy. Recently, we have shown that heteromers formed by the dopamine D2 and serotonin 5-HT1A receptors (D2R and 5-HT1AR, respectively) might be important site of action of clozapine 7. Using the conventional route of administration, this drug (as well as other antipsychotics), addressed only for a small part of the cell population in the brain, is non-specifically delivered into all brain areas, causing undesirable side effects. Higher selectivity may be achieved using compounds directly targeting D2-5-HT1A heteromers because these heteromers can be formed only on neurons co-expressing receptors engaged in the complex formation.
Human monoclonal scFv (single-chain variable fragment) antibodies may be used as targeting ligands to decorate the outer surface of nanovehicles. ScFvs are small antibody fragments that compose the variable regions of the heavy (VH) and light (VL) chains of immunoglobulins with a flexible peptide linker designed to connect the two chains in such a way that the antigen binding site is retained in a single co-linear molecule 8. ScFvs can be obtained from phage display libraries 9, 10. ScFvs have the potential to be very useful for the targeted delivery of drugs to specific cells or tissues. In comparison to the much larger Fab, F(ab)2, and IgG forms of monoclonal antibodies, from which they are derived, scFvs have lower retention times in non-target tissues, faster blood clearance, better tissue penetration and reduced immunogenicity, making them attractive for therapeutic applications 11.
Therefore, in the present study, we focused on the production of an scFv antibody that specifically binds the D2-5-HT1A heteromer. The antibody should be able to recognize the heteromer formed by both receptors, and it should not recognize the monomeric forms of the receptors. The obtained antibody would be used to modulate the surface properties of nanocapsules, which could then be exploited as a novel nanocarrier of antipsychotics. In this way, we can obtain efficient delivery of the encapsulated drugs preferentially to the defined site in selective tissues. To produce specific scFv antibodies with defined a D2-5-HT1A heteromer selectivity, the antibody phage display libraries Tomlinson I+J was used. The library allows us to obtain approximately 3x108 various filamentous phages with the relevant fusion protein PIII-scFv (coding by phagemid pIT2) as part of their coat. The bio-panning process was conducted in a CHO-K1 cell line expressing the desired receptors.
Materials and Methods
Cell Culture and Transfection
CHO-K1 cells (ATCC) were grown in RPMI (Sigma) medium supplemented with 10% heat-inactivated fetal bovine serum (FBS) (Sigma). The TurboFect reagent (Thermo Sci.) was used for transient and stable transfections. The procedure was carried out in accordance with the manufacturer’s protocol. Early passages of CHO-K1 cells were stably transfected (1.5 mg DNA) with the plasmid pcDNA3.1(+) encoding the human 5-HT1AR or the human D2R (UMR cDNA Resource Centre) separately or cotransfected with both vectors. Stable cell lines expressing D2R and/or 5-HT1AR were obtained after the addition of the selection antibiotic, G418 (Sigma), at a final concentration of 0.75 mg/ml. Cells resistant to the antibiotic and stably expressing investigated receptors were analysed by RT-PCR (data not shown). Forty-eight hours before the selection experiment, stable cell lines were transiently transfected with 0.5 mg DNA (per 10 mm plate area) encoding the desired receptors. Additionally, the HEK-293 cell line (ATCC) was used. The cells were grown in MEM (Sigma) supplemented with 1% L-glutamine and 10% heat-inactivated FBS (Sigma).
All cells were cultured at 37°C inside a humidified incubator in an atmosphere of 5% CO2.
Screening of Phage-Displayed scFv Libraries
The human antibody scFv phagemid library Tomlinson I+J (Geneservice) was used. The library J was amplified and titrated according to the manufacturer’s protocols using E. coli TG1 cells. Biopanning was conducted against the D2-5-HT1A heteromer using CHO-K1 cells expressing D2-5-HT1A heteromers(CHO-positive cells; CHO+) and CHO-K1 cells expressing D2R mixed with CHO-K1 cells expressing 5-HT1AR (1:1) (CHO-negative cells; CHO-). Five rounds of positive selection followed by negative preselection and finally 2 negative selections were performed. Preselection was performed as follows: amplified phages were blocked for 2 hr at RT (or overnight at 4°C) in 3% MPBS and then added to culture medium with CHO- cells grown on 150 mm plates (20x, 95% confluence) and incubated for 2 hr at 0°C. Then, the temperature was changed to 37°C for 20 min. The CHO- cells were centrifuged (10 min at 1000 rpm), and the supernatant containing unbound phages was added to the medium with CHO+ cells grown on 150 mm plates (10x, 95% confluence). From that moment, the positive selection began and extended for 2 hr at 0°C. In the next step, the cells were washed 4 times with cold PBS. Between washings, RPMI medium was added, and the cells were incubated on ice for 10 min. Similar to preselection, for the next 20 min, the temperature of incubation was changed to 37°C. In the next step, the cells were washed 4 times using elution buffer (100 mM glycine, 150 mM NaCl, pH 2.8). The number of washes increased (twice each time) with subsequent rounds of selection. Finally, CHO+ cells were harvested from the plates and resuspended in PBS containing trypsin (1 mg/ml) for approximately 15 min (until cell lysis). Then, the obtained suspension was centrifuged (10 min, 1000 rpm, 4°C), and the supernatant containing the desired phages after titration and amplification was used for another round of biopanning. The biopanning rounds were repeated until the results of polyclonal phages ELISA related to output phages reached a plateau or started to decline.
Polyclonal Phage ELISA
The efficiency of the biopanning process was determined using polyclonal phage ELISA. A total of 50 ml of amplificated phages (obtained after each round of selection) was added to each well of the 96-well V-microtiter plate (Sarstedt) and incubated with 50 ml of 4% MPBS for 2 hr at 37°C. Then, 1.5 x105 CHO+ cells (resuspended in 50 ml of RPMI with 5% FBS) expressing D2-5-HT1A heteromers (positive probe) were added to the wells with blocked phages and incubated on ice for 1 hr. The negative probe consisted of CHO- cells, i.e., cells expressing a single type of receptor mixed at the 1:1 ratio. The washing step was conducted 3 times at 4°C using 200 ml cold PBS. After each round, the cells were centrifuged (300 x g, 10 min, 4°C), and the supernatant was rejected. Bound phages were detected using horseradish peroxidase (HRP)-conjugated anti-M13 monoclonal antibodies (GE Healthcare). Briefly, after washing, the cells were incubated with the antibody resuspended in a 1:5000 ratio in 3% MPBS for 30 min on ice and then washed 4 times as described above. Finally, 100 ml of TMB substrate (GE Healthcare) was added to each well. The reaction was stopped with 1 M HCl (100 ml per well), and the absorbance was measured at 450 nm. Experiments were performed in triplicate.
Monoclonal Phage ELISA
Based on the results of polyclonal phage ELISA, phage clones from rounds characterized by the highest affinity against CHO+ cells were randomly selected for monoclonal phage ELISA experiments. Individual colonies were inoculated into 96-well plates containing 100 ml 2xTYAG (2xTY (Bioshop) with 100 μg/ml ampicillin (Sigma) and 1% glucose (Bioshop)) medium per well and cultured overnight at 37°C (250 rpm). Then, 5 ml of the culture (from each well) was added to fresh 200 ml 2xTYAG medium and cultured with shaking (250 rpm) at 37°C for 2 hr. After that time, 109 helper phages were added to the wells and incubated for 1 hr and at 37°C with shaking at 250 rpm. The plates were then centrifuged (1800 x g, 10 min), the supernatants were aspirated off, and bacterial pellets were resuspended in 200 ml 2xTYAKG (2xTY containing 100 μg/ml ampicillin, 50 mg/ml kanamycin (Sigma) and 1% glucose) medium and incubated overnight at 30°C (250 rpm). Finally, the plates were centrifuged (1800 x g, 10 min), and 50 ml of supernatants (containing monoclonal phages) was used in the phage ELISA as described above (2.4).
ScFv Expression in the Bacterial System
To obtain soluble scFv expressed in E. coli HB2151, a single colony was used to inoculate 5 ml of 2xTY medium containing 100 μg/ml ampicilin, and the cultures were grown overnight at 37°C. A total of 2.5 ml of the overnight culture was used to inoculate 25 ml, and then this culture was used to start a 1 L culture at 37°C. The expression was induced in the host bacteria by adding IPTG to the final concentration of 1 mM when the OD600 reached 0.9; the incubation continued for 16 h at 28°C, and then the cells were harvested by centrifugation (4000 x g, 15 min, 4°C). The cells were placed on ice until scFv purification.
Large-Scale Purification of Soluble scFv Fragments
ScFvs are secreted into the periplasmic space located between the outer and inner membrane of E. coli. The pellet of bacterial cells was resuspended in buffer consisting of 50 mM Tris-HCl (Bioshop), pH 8.0; 20% sucrose (Bioshop); 1 mM EDTA (Bioshop); and 1x Protease Inhibitor Cocktail (Sigma) using 20 ml buffer/1 L overnight culture). The cells were incubated on ice for 15 min with gentle agitation, and then the cell suspension was centrifuged at 30000 x g for 30 min at 4°C. The supernatant is the osmotic shock fluid containing periplasmic proteins and was divided into two equal parts: one was dialyzed extensively against Protein L Binding Buffer (100 NaH2PO4, pH 7.2; 150 mM NaCl) (Bioshop). The dialysis was carried out at 4°C overnight before continuing with the purification. The supernatants were centrifuged (30000 x g, 30 min, 4°C) after dialysis. ScFvs were purified at room temperature.
The extract of E. coli scFvs was loaded onto a column filled with 5 ml of Protein L Agarose equilibrated with Protein L Binding Buffer at a flow rate of 1 ml/min. After washing with 10 column volumes of Binding Buffer, the proteins were eluted from the column with Elution Buffer that contained 0.1 M glycine, pH 2.5 (at flow rate 1 ml/min). Eluted fractions were immediately adjusted to physiologic pH by adding 100 μl of the Neutralization Buffer (1 M Tris, pH 8.0) to 1 ml of eluate. Fractions containing scFvs (as judged by SDS-PAGE and Coomassie Brilliant Blue staining) were pooled and dialyzed against 1x PBS at 4°C overnight. The concentration of protein was calculated via Bradford assay. The purified fractions were stored at -80°C.
SDS-PAGE
Samples for analysis were mixed with 2x Electrophoresis Buffer containing 50 mM Tris-HCl, pH 6.8; 10% glycerol; 2% SDS; 0.1% bromophenol blue and 5% β-mercaptoethanol and heated for 5 min at 95°C. SDS-PAGE was performed according to Laemmli, 1970 12 with a 4% stacking gel and a 12% resolving gel (Mini-Protean 3, Bio-Rad). Gels were stained by Coomassie Brilliant Blue.
Analysis of Specificity of Obtained scFvs Monoclonal Antibodies
The specificity of purified scFvs was determined by ELISA and flow cytometry analysis. In the case of the ELISA experiments, 100 ml of purified antibodies at different concentrations, resuspended in 4% BSA, were added to wells of a 96-V-well plate and incubated for 1.5 h at 37°C. Then, 1.5 x105 CHO+ cells as well as CHO- cells (resuspended in 50 ml of RPMI with 5% FBS) were added to the suitable well with blocked antibodies and incubated on ice for 1 hr. The washing step was conducted 3 times at 4°C using 200 ml cold PBS. After each round, the cells were centrifuged (300 x g, 10 min, 4°C), and the supernatant was aspirated off. Then, the pellets were resuspended in 50 ml protein L conjugated with HRP (protein L was resuspended in 0.5% BSA/PBS, 1:2000 ratio) and incubated for 30 min on ice. After washing (carried out 4 times as described above), the reactions were developed by TMB substrate. Experiments were performed in triplicate.
Moreover, similar experiments were conducted with HEK-293 cells. HEK-293+ represents cells stably expressing D2R and 5-HT1AR, HEK-293- represents a mixture of HEK-293 cells expressing individual forms of the receptors.
Before experiments using flow cytometry analysis, purified antibodies were conjugated with fluorescent dye. The idea of the tests was the same as described above in the case of ELISA. The same types of cells were used. The experiments were conducted to confirm the results from ELISA.
Results and Discussion
Phage display is a powerful tool for the identification of peptides, proteins or antibodies with affinity for a specific target 13. The advantage of the method is the possibility of the production of monoclonal antibodies recognizing antigens that cannot be used to immunize an animal due to their toxicity, non-immunogenicity or presence in complexes on the surface of cell membranes 14. Therefore, in the present study, we used a phage display technique to prepare a novel human monoclonal scFv antibody that specifically recognized the D2-5-HT1A heteromer. To obtain such properties, the antibody should recognize the structural, spatial epitope formed within the heteromer structure. Both receptors mentioned above are membrane proteins belonging to the GPCR family. Therefore, it was impossible to perform a selection process using recombinant antigen since obtaining such an antigen in a soluble form with the correct spatial structure is too difficult to achieve in the case of membrane proteins.
In the present study, to separate phages specifically binding to the D2-5-HT1A heteromer, the selection process using CHO+ cells with overexpression of both investigated receptors was conducted. Phages that were bound to receptor monomers or other CHO-K1 cell surface proteins were eliminated as a result of negative selection by using CHO- cells expressing separate receptor monomers. To increase the density of receptors on the surface of CHO-K1 cells, the stable cell lines were additionally transiently transfected with plasmids encoding receptors of interest. The cells of the CHO-K1 cell line are very useful in such experiments since they are easy to culture and are susceptible to transfection; they do not express D2R or 5-HT1AR endogenously, which is a very important issue because it provides the opportunity to modulate the density of the receptors on the cell surface. The biopanning process was based on library phages interacting with the antigen formed on the cell surface by the D2-5-HT1A heteromer. The process was performed on cells growing on the plate surface. The process provided the best conditions for the formation of the spatial D2-5-HT1A antigen and minimized the risk of isolation of antibodies that recognize undesired antigens 15. Several parameters affect biopanning efficiency, including antigen concentration, temperature, washing stringency (washing number and composition of wash buffer) as well as blocking and elution buffer composition 16.
Elimination of nonspecific binding without losing rare specific clones remains a serious challenge 16. Therefore, in the present study, we decided to perform negative preselection in each round of biopanning. Finally, after 5 full rounds (preselection and positive selection), 2 negative rounds were conducted. During each round, after incubation at 0°C, the temperature was changed to 37°C. It induces the internalization process. In our previous studies, the different kinds of selections were tested, and the highest level of phage enrichment was observed in the case of isolation of phages after the internalization of receptors serving as the desired antigen (unpublished results).
The selection process was assessed by monitoring the enrichment ratio and polyclonal phage ELISA. The increasing titre of phages indicates the correctness of the bio-panning process and corresponds with the enrichment of phages that specifically recognized the D2-5-HT1A heteromer (Figure 1). In the initial rounds, the difference in polyclonal phages binding to CHO+ vs. CHO- cells was small, while after the 4th and 5th round, the positive signal was more than 8 times higher.
Figure 1. Monitoring of bio-panning process. A) Titre of phages after each selection rounds; B) Enrichment of phages that specifically recognized the D2-5-HT1A heteromer – polyclonal phage ELISA.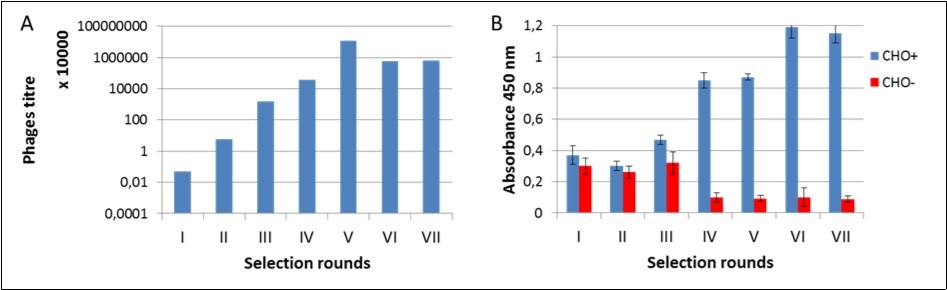
After the selection process, the specific binding of individual monoclonal phages to CHO+ cells was tested by ELISA techniques. Experiments were performed using CHO+, CHO- and CHO-K1 cells. Twenty-six phages of 960 tested exhibited high levels of D2-5-HT1A heteromer specificity (Table 1).
Table 1. Binding level of 12 various monoclonal phages specific to D2-5HT1A heteromer (CHO+ cells) in relation to: CHO-K1, CHO- cells – determined by ELISA technique.| scFv name | CHO-K1R | CHO-R |
| 1F/5r1 | 3.87 | 3.62 |
| 4C/5r1 | 3.67 | 3.35 |
| 5E/5r1 | 4.44 | 4.67 |
| 1G/5r1 | 46.21 | 41.83 |
| 10G/5r2 | 18.81 | 16.82 |
| 4H/5r2 | 2.83 | 2.56 |
| 6H/5r2 | 16.87 | 16.32 |
| 4D/5r3 | 2.23 | 2.31 |
| 5D/5r3 | 2.98 | 2.27 |
| 8H/5r3 | 4.21 | 4.32 |
| 2D/5r4 | 22.57 | 23.86 |
| 6C/5r5 | 5.27 | 4.37 |
Among originally selected phages, satisfactory expression could be obtained only for 12 clones. This was due to the presence of STOP codons within the sequence of DNA encoding the antibodies (data not shown). Four phages possessing the best parameters were used for the infection of bacterial cells HB2151 (E. coli) and the production of soluble monoclonal scFv antibodies. The expression level was monitored by SDS PAGE electrophoresis. The antibodies were obtained from periplasmic extract and purified by affinity chromatography using Protein L-immobilized resin.
The activity of phage display-derived antibodies was determined using ELISA (Figure 2). The specificity of D2-5-HT1A heteromer recognition of 4 soluble scFv fragments was tested using CHO+, CHO-, CHO-K1 cells as well as CHO-K1 cells expressing different GPCR - somatostatin receptor type 1 (Sst1). Additionally, similar tests were conducted using HEK-293+, HEK-293-, and HEK-293 cells. Similar results were recorded using flow cytometry techniques (data not shown). Based on the results, the 1H/5r1 antibody exhibits the most promising properties.
Figure 2. Phage display-derived scFv antibodies. A) SDS-PAGE electrophoresis after protein purification; B) D2-5-HT1A heteromer specificity of isolated scFvs antibodies determined using ELISA technique (y axis – ratio of positive vs negative signal).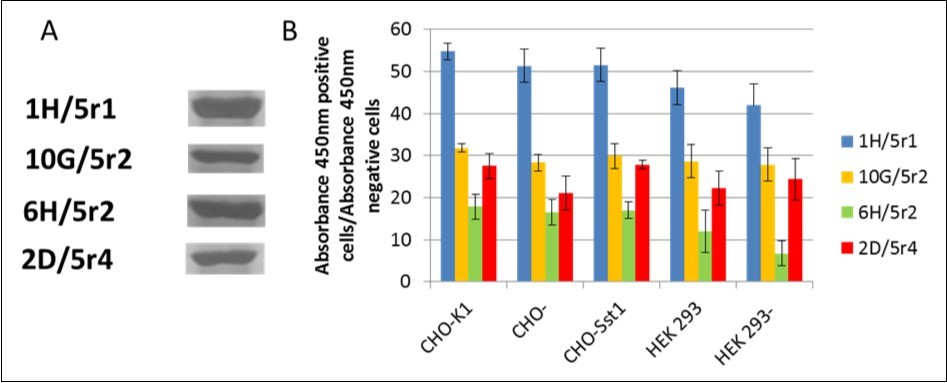
In conclusion, it has been shown here that phage display methodology can be successfully used in the preparation of monoclonal antibodies against membrane proteins. In the present study, a novel human monoclonal scFv antibody, which specifically recognized the heteromer formed by D2R and 5-HT1AR and simultaneously did not bind to the monomeric forms of the receptors, was isolated. In the future, to obtain the best parameters of antigen-antibody interaction, the modifications of selected antibodies will be conducted using random mutagenesis. Eventually, the obtained antibody would be used as a targeting ligand to functionalize the surface of nanocapsules, which would then be exploited as a novel nanocarrier of antipsychotics.
Acknowledgments
The manuscript has been corrected by AJE - Certificate Verification Key 7E03-CDAE-8EC3-6773-9BB7. The Faculty of Biochemistry, Biophysics and Biotechnology is partner with the Leading National Research Centre (KNOW) supported by the Ministry of Science and Higher Education. This work was supported by grants 35+ Nr. 35p/1/2016 from KNOW. The work was also co-financed from European Union within Regional Development Fund - Grants for innovation – PARENT/BRIDGE Programme - POMOST/2011-4/5.
Conflict of Interest
The authors report no conflicts of interest in this work. The authors also declare no competing financial interest.