Abstract
We explored the effect of a change in substrate-benzoate (as sole carbon and energy source) concentration in growth medium on the activity of benzoate 1,2-dioxygenase (BDO) of R.opacus 1CP cells, where BDO is the enzyme mediated the initial attack of benzoate. The activity of the enzyme was estimated by a change of respiration of whole freshly harvested bacterial cells (growth of the cells on benzoate) in response to injection of benzoate. It was shown that when concentration of growth substrate-benzoate decreased from 6 g/L to 250 mg/L, the curves of the dependency of the response rate to benzoate on the initial concentration of benzoate demonstrated that kinetics of the process changes from hyperbolic saturation kinetics, or typical Michaelis-Menten kinetics, to sigmoidal dependency of V on S. The semisaturation constant as a characteristic of the strength of substrate binding with BDO changed simultaneously. These changes were accompanied by the increase in the Hill coefficient from 1.02 up to 3.06, hence positive kinetic cooperativity by a substrate was observed for BDO of R.opacus 1CP cells. The influence of this type of cooperativity on viability of rhodococcus in natural environment and causes of the changes mentioned are discussed. It was hypothesized that an increase in substrate concentration in the medium for the growth of the bacterium not only stimulated synthesis of the inducible enzyme (BDO) in the cell but also led to the change in BDO conformation followed by the change in interaction between substrate-binding active sites of enzymes.
Author Contributions
Academic Editor: Syed Shams ul Hassan, Ocean college Zhejiang University
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2017 Elena V. Emelyanova, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Competing interests
The authors have declared that no competing interests exist.
Citation:
Introduction
Under aerobic conditions the initial attack of different substrates by bacteria is usually performed by mono- or dioxygenases 1. In many cases these enzymes are complex systems. Benzoate degradation under aerobic conditions starts from an initial attack of this compound by benzoate 1,2-dioxygenase (BDO). BDO is a two-component enzyme consisting of oxygenase and reductase components, and as example, oxygenase component has an (alpha beta)(3) subunit structure 2.There are some difficulties in determining BDO activity in a cell-free extract 3, 4. For instance, the determination of the BDO activity in a cell-free extract of P. arvilla C-1 required introduction of NADH-cytochrome c reductase, but even in that case the BDO activity was just 6% of the activity at the reductase saturation 5.The BDO activity in a cell-free extract could not always be determined even if the depletion of substrate was registered 6. In majority of cases, the rate of BDO reaction is determined in whole cells by monitoring their respiration in the presence of respiration substrate 7. We have shown that the change of freshly harvested cells respiration in the presence of benzoate was an indicator of BDO activity of R.opacus 1CP cells 8. While studying growth of R. opacus1CP on benzoate as a sole source of carbon we found that not only growth characteristics of the culture but also a kinetics of enzymatic reactions and the enzyme pool of the cell were changing when the benzoate concentration in the cultivation medium was raised 9, 10. It was found that a plot of change of oxygen consumption by cells versus benzoate concentration deviated from the Michaelis-Menten hyperbolic dependency and had an S-shaped form when the cells grew on benzoate at low benzoate concentration. A sigmoidal but not Michaelis-Menten hyperbolic plot of the dependency of the rate of enzymatic reaction on the initial substrate concentration is typical for allosteric enzymes.
It is known that at least one allosteric enzyme providing regulation of total reaction of whole enzyme ensemble is present in every multienzyme complex. More often this enzyme catalyzes the rate of the first (as it is in the case of BDO and benzoate) and the slowest reaction 11. The presence of several active and some allosteric centers is a characteristic feature of an allosteric enzyme molecule 12. Allosteric enzymes possess unusual kinetic properties, one of which markedly exerts in S-shape type of a plot 13, 14. At the same time not any kinetic anomaly such as S-shape type of the plot is of allosteric nature. Allosteric interactions are also possible for monomeric enzymes 15. The statement concerning allosteric nature of kinetic anomalies in the behavior of the enzyme is valid only when the presence of a subunit structure and the existence of several substrate binding centers in the enzyme are proven 13, 14.
The aim of the present study was to compare kinetic constants, obtained for BDO of R. opacus1CP cells, grown under different benzoate concentrations and at first time to find a connection between them and cooperativity by a substrate.
Materials and Methods
The Microorganism and Culture Conditions
The object of our study was Gram-positive nonspore-forming bacterium Rhodococcus opacus 1CP isolated from the selective medium with 2,4-dichlorophenol and able to grow on media with aromatic compounds including benzoate 16.
The strain has been stored as resting cells at +4ºC in a sterile buffer for 5 years. For cell activation and inoculum preparation 10 μL of resting cell suspension were inoculated on enriched agarized Luria-Bertani (LB) medium and incubated at 28ºC until germination. Germinated cells were washed out with 10 mL of mineral medium (see below) and inoculated on agarized mineral medium with benzoate (200 mg/L) as a sole growth substrate.
To produce biomass, R. opacus 1 CP was grown on benzoate-containing mineral medium of the following composition (g/L): Na2HPO4 – 0.73; KH2PO4 – 0.35; MgSO4´7H2O – 0.1; NaHCO3 – 0.25; MnSO4 – 0.002; NH4NO3 – 0.75; FeSO4´7H2O – 0.02. Biomass grown on Petri dish with solid mineral-benzoate medium was washed out with mineral medium and used as an inoculum. The culture was incubated in 750-mL Erlenmeyer flasks containing 100-200 mL of the abovementioned medium with benzoate at a concentration from 250 mg/L to 6 g/L (28ºC, 220 rpm).
Determination of BDO Activity of R. opacus 1 CP Cells
BDO activity of R. opacus 1 CP cells was determined as the rate of respiration change of suspended freshly harvested cells in the presence of benzoate. To perform determination, R. opacus 1 CP cells were grown on benzoate to an optical density of 0.5–2.2 (595 nm), centrifuged (12,000-16,000 g, 10-15 min, 4ºC), washed twice with 50-mM Tris-HCl, pH 7.2, suspended in the same buffer and immediately used for analysis. The measurements were carried out in air-saturated 50-mM Tris-HCl buffer, pH 7.2 at room temperature in an open 5 mL cuvette equipped with a mixer. To determine BDO activity, after base level of cell respiration was stabilized, benzoate was injected into the cuvette, and the rate of oxygen concentration change was measured with a Clark’s oxygen electrode. The oxygen electrode was equipped with an Ingold 531 O2 amplifier (Switzerland-USA). The signal was recorded with a two-coordinate XY Recorder-4103 (Czech Republic). The recorded signal reflected the rate of the enzymatic reaction of BDO with substrate. The unit used to measure rate was pA/s.
Experimental Data Processing
SigmaPlot program (version 12) was used to calculate the constants of Hill, Michaelis-Menten equations and plot linear regression curves using experimental data on R.opacus 1CP BDO activity obtained for cells grown in medium with different benzoate concentration.
Statistics
The measurements were taken in triplicate in two independent series of experiments. The presented results reflected averaged values. Statistical analysis of the data was carried out using a Student’s t-test, taking as sufficient the probability criterion P < 0.05.
Results
R.opacus 1CP cells were grown in benzoate-containing medium, where benzoate concentration varied from low to high: 250 mg/L - 6 g/L. Complete degradation of benzoate at concentration of 250 mg/L occurred for less than a day, maximum density of the culture achieved 0.5 optical units of density (OD) at λ = 595 nm. It took up to 65 h for the culture to degrade 6 g/L of benzoate, OD was 2.2 units. During cultivation a lag-phase did not exceed 3 h and 15 h at benzoate concentration of 250 mg/L and 6 g/L, respectively.
Freshly harvested cells were suspended in Tris-HCl buffer, BDO activity of cells was determined and the curves of the dependency of the rate of cell reaction catalyzed by BDO of R.opacus 1CP cells on the initial benzoate concentration were plotted. The curves of the reaction to benzoate which were plotted for the cells grown in the medium with 250 mg/L, 2 and 6 g/L of benzoate are given in Figure 1. The curves (rate vs substrate concentration) for the whole interval of the initial benzoate concentrations are represented in Figure 1a. To better visualize the differences in these curves, the area of low benzoate concentrations is shown in Figure 1bin larger scale.
Figure 1. A plot of the rate of BDO reaction (V) versus the initial concentration (S0) of substrate-benzoate (a – all tested benzoate concentrations; b - the area of low benzoate concentrations) for R. opacus 1CP cells grown at different benzoate concentrations: 250 mg/L, 2 and 6 g/L.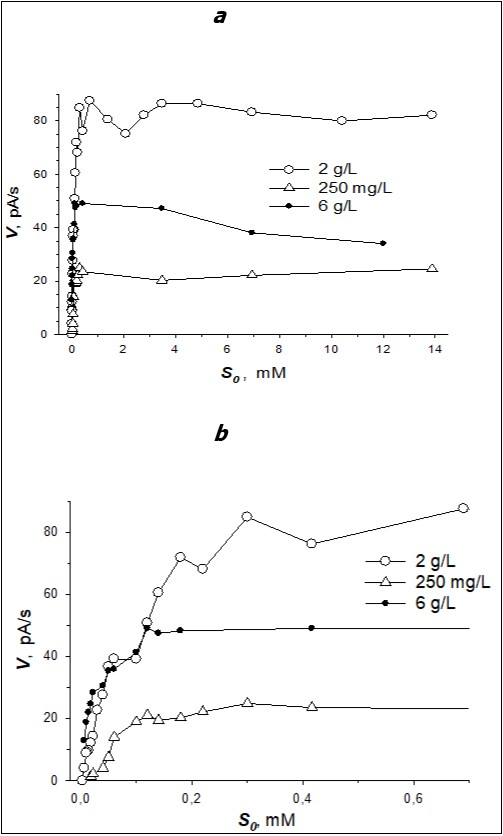
Equations of nonlinear regression such as Vmax·Sn/ (S0.5n + Sn) were calculated based on experimental data using SigmaPlot, where Vmax is maximum rate of reaction catalysed by enzyme (i.e. maximum value of the rate when S→∞); S0,5 is substrate (S) concentration at which V=0.5Vmax; n is the Hill coefficient. For R.opacus 1CP cells, grown at different benzoate concentrations in growth medium, the kinetic constants of BDO-catalyzed rate of cell response to benzoate were obtained. These constants are summarized in Table 1.
Table 1. Coefficients of the Hill equation calculated for the reaction catalyzed by BDO of R. opacus 1CP cells grown on benzoate at different concentrations| Benzoate concentration ingrowth medium, mg/L | S0.5,mM | Vmax,pA/s | n |
| 250 | 0.058 | 22.6 | 3.06 |
| 2000 | 0.069 | 84.0 | 1.32 |
| 6000 | 0.019 | 50.74 | 1.02 |
When the cells were grown in the medium at benzoate concentration less than 6 g/l, the calculated the Hill coefficients (n) were not equal to 1. It points to the fact that the dependency of V on S for BDO of cells grown at those concentrations of benzoate was not hyperbolic. To verify the sigmoidal dependency, experimental data are depicted with reciprocal coordinate 1/V-1/S (Figure 2a-c).
Figure 2. A plot of the rate of the reaction (V, pA/s) catalysed by BDO of R. opacus 1CP cells versus the initial concentration of substrate-benzoate (S0, mM) in reciprocal coordinates 1/V-1/S (dots: experimental data; line: linear curve) for the culture grown in medium with benzoate concentrations: a – 250 mg/L; b - 2 g/L; c – 6 g/L.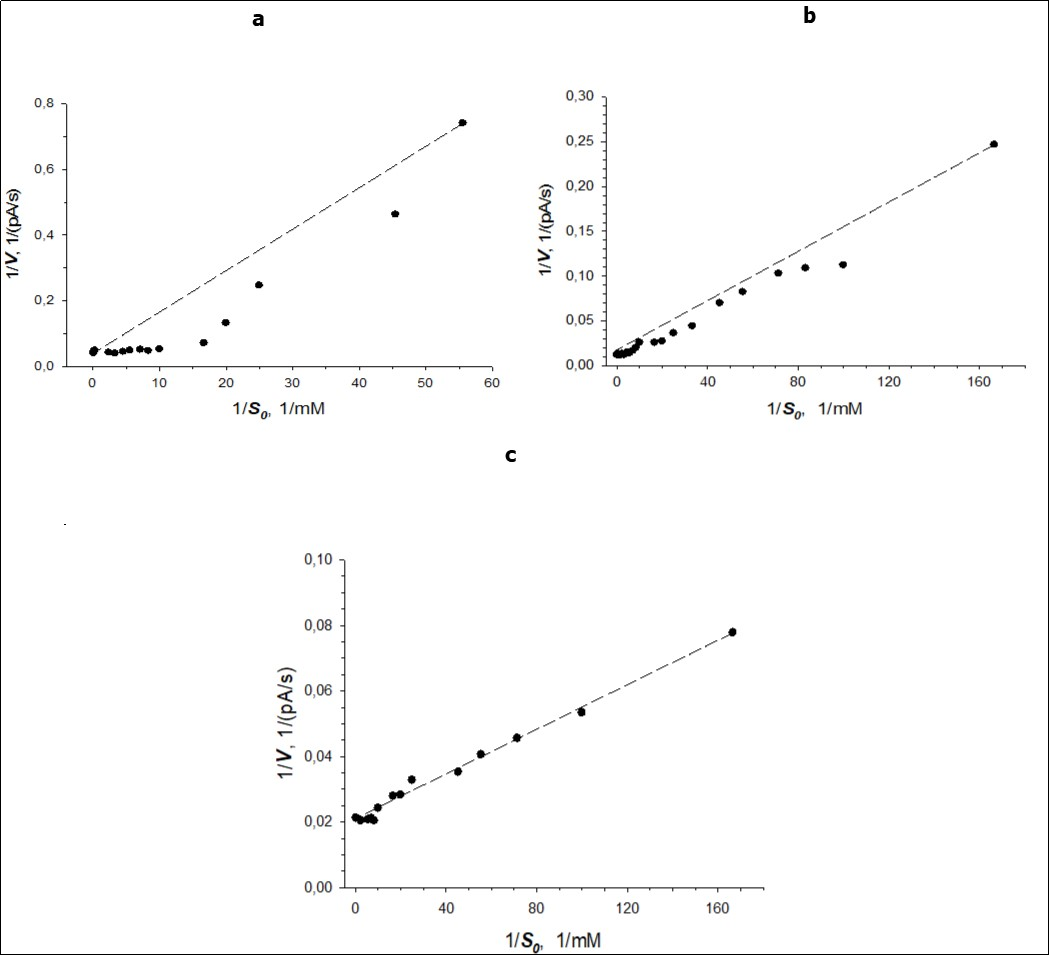
Discussion
The change of 1CP respiration in response to benzoate injection into cell suspension is a combined response to benzoate of the whole cell containing BDO. M. Dixon and E. Webb 17 wrote that enzymes had been divided into two groups as "unorganized" and "organized" and this approach has been used until recently 18. Taking into account modern understanding of the subject the enzymes are grouped into extracted enzymes and enzymes functioning only in the live cell. Benzoate 1,2-dioxygenase is among the enzymes the activity of which can be fully estimated in the live cell. Earlier we have shown that the response of freshly harvested cells (benzoate is growth substrate) of R. opacus 1CP to benzoate is an indicator of the activity of benzoate 1,2-dioxygenase. Therefore, in this study the change of respiration of freshly harvested cells in response to benzoate was considered as a parameter characterizing the activity of BDO in this culture.
Using the cells grown at benzoate concentration of 6 g/L it was found that the dependency of the rate of enzymatic reaction on the initial benzoate concentration could be described with equal certainty both by hyperbolic Michaelis-Menten equation and Hill equation:
When the cellsof R. opacus 1CP were grown in the medium containing benzoate at concentration of less than 6 g/L, the dependency of V on S for BDO of cells was not hyperbolic: for these cells the dependency of 1/V on 1/S was not linear (Figure 2). Sigmoidal curves (V vs S) were obtained. Kinetic constants determined for BDO ofbacterial cells, represented in Table 1, support this.
Obtained sigmoidal curves can be described using empirical formula suggested by Hill:
Michaelis constant, Km, more often determined by graphical means as Km=S0,5, called the apparentMichaelis constant, is widely used for characterization of the strength of substrate binding with the enzyme. However, it is not a true constant of dissociation of an enzyme-substrate complex(Km). This is a concentration of a substrate at which the rate is equal to half past maximum (V = 0.5Vmax). Hence in a case of R.opacus BDO, we think that S0,5 represented in Table 1, can be used for estimation of the strength of substrate binding with BDO.
Table shows that an increase in benzoate concentration in the medium of bacterium cultivation led not only to deviation of the curves (V vs S) from simple Michaelis-Menten kinetics but also had an effect on the strength of substrate binding with BDO.
A molecule of the allosteric enzyme contains multiple active and regulatory sites. Allosteric interactions are found by deviation of the curves V vs S from typical Michaelis-Menten kinetics 19.
In our case for BDO of R. opacus 1CP cells, grown in the medium containing 250 mg/L and 2 g/L of benzoate (Figure 1), deviations from hyperbolic kinetics were diverse. Sigmoidal curves of the dependency of V on S which had more than one flex points and intermediate plateau were obtained. A sharp turn of the curve forming a plateau was observed.
The Hill coefficient (n) is a dimensionless parameter. It may characterize the number of substrate-binding active sites 20. n is used as a parameter summarizing the degree of cooperatives 19. The Hill coefficient is > 1 under “positive kinetic cooperativity” by a substrate and < 1 under “negative kinetic cooperativity”. S-shape curve was obtained while studying glutamate dehydrogenase 21. Positive cooperativity was observed.
Concavity of a curve of 1/V-1/S(Figure 2) for BDO of R. opacus 1CP cellspoints to “positive kinetic cooperativity” by a substrate.
The Hill coefficient (3.06) for BDO of cells, grown in the medium containing 250 mg/L of benzoate also indicates the presence of positive cooperativity. n decreased up to 1 when benzoate concentration in the growing medium increased. As shown in Figure 2 concavity of the curves of 1/V-1/S disappeared, when the cells were grown at benzoate concentration of 6 g/L.
An increase in the Hill coefficient as positive kinetic cooperativity by substrate for allosteric enzymes as assumed in 13, 14 is caused by interaction between sites binding a substrate. For allosteric enzymes, in order to prove spatial disconnectedness between sites a desensitization approach is employed 19, 22. Desensitization can be induced by physical impact or by chemical modification. After desensitization enzymatic activity is not lost.
To confirm the allosteric nature of positive kinetic cooperativity by a substrate the approaches used for desensitization of allosteric enzymes are helpful. For instance, heat (60°C for 4 min) desensitization of the allosteric enzyme of E. coli was accompanied with the increase in specific enzymatic activity and disappearance of cooperativity by a substrate 23, 24.
If after desensibilization S-shape plot of the dependency of the rate on the initial substrate concentration alters to hyperbolic plot, then it is the evidence of the allosteric nature of the kinetic cooperativity by a substrate.
However, we used a response to benzoate of the whole cells containing BDO in order to estimate the activity of BDO of R. opacus 1CP cells. That is why we could not apply such action on the cell as heating or chemical modification that led to the change in the reaction not only of the enzyme but also of the whole cell.
Positive kinetic cooperativity is typical for the enzymes catalyzing stages which limit the general rate of metabolism (BDO is the enzyme mediated the initial attack of benzoate). Due to this cooperativity the range of substrate concentrations necessary for substantial alteration in the activity of the enzyme narrows: the activity of the enzyme becomes more sensitive to the change in substrate concentration. As a consequence, the semisaturation point was observed at lower substrate concentration (Figure 1b). Rhodococci are representatives of dispersion microflora. These microorganisms adopted to live at low substrate concentrations. The physiological significance of positive kinetic cooperativity for rhodococci lies in the fact that it is sufficient to have traces of a substrate to activate metabolism. Moreover, saturation is achieved at lower substrate concentrations; this is important for bacteria, for which toxic substances often serve as substrates. In this case positive kinetic cooperativity by a substrate plays a protective role. In the cell an excess of substances which are dangerous for a microorganism is undesirable. Hence, it is important that activation of metabolism takes place at low substrate concentrations. The process of substance transport into the cell and its metabolism are interrelated, inhibition of the activity of the enzyme by not high concentration of dangerous substrate prevents further transport of the substance into the cell.
Allosteric enzymes are oligomers consisting of identical protomers. So, benzoate 1,2-dioxygenase of Pseudomonas putida mt-2 2 is an oligomer with an odd number of protomers. Displacement of equilibrium between oligomer forms of the allosteric enzyme may lead to the deviation from typical Michaelis-Menten kinetics, the appearance of sigmoidal curves and kinetic cooperativity by a substrate. For some enzymes a hyperbolic dependency of the rate of enzymatic reaction on the initial substrate concentration changed to a sigmoidal dependency at rather high substrate concentrations 25. For other enzymes a change from hyperbolic to S-shape was observed when enzyme concentration decreased 26.
It was impossible to determine specific activity of BDO of R.opacus 1CP cells on the basis of estimated cell response to benzoate. However, induction of the BDO activity in the cells without BDO at benzoate concentration of 6 g/L occurred more intensively compared to that at benzoate concentration of 300 mg/L (data not published). Using these indirect data it can be assumed that the content of the enzyme in the cells grown in the medium at benzoate concentration of 250 mg/L was lower than that during the growth at benzoate concentration of 2 g/L. In parallel, the curves became less sigmoidal when the benzoate concentration was changed from 250 mg/L to 2 g/L.
The allosteric enzymes consisting of several subunits can be dissociated into separate subunits with other catalytic properties than the initial oligomer or associated. The change in the dissociation degree of the enzyme molecule may occur both in the change in enzyme concentration and under the action of a substrate. Under dissociation-association of the enzyme the enzymatic activity may disappear.
For BDO of R. opacus 1CP cells when concentration of substrate-benzoate was elevated in the growth medium, no loss of the activity was observed, but a decrease and disappearance of cooperativity by a substrate were observed.
When substrate concentration was increased in the growth medium, probably, not only the inducible enzyme (BDO) was induced, but its conformation was also changed leading to the alteration in interaction between substrate-binding active sites. Consequently, the sigmoidal dependency of V on S disappeared and the curve of the dependency of the rate of enzymatic reaction on the initial substrate concentration obeyed the hyperbolic Michaelis-Menten kinetics.
Conclusion
Thus, peculiarities, determined for BDO of R.opacus 1CP cells during the change of benzoate concentration in the medium where the culture grows and in its cells the enzyme is induced, are explained, first of all, by the allosteric nature of the enzyme.
Further treatment without damage (for instance, storing at 0°C) of the cells grown at benzoate concentration of 250 mg/L-4g/L will be applied to confirm the allosteric nature of kinetic cooperativity by a substrate for BDO of R. opacus 1CP cells. In case when S shape of the curves disappears, the allosteric nature of the mentioned anomalies is confirmed.