
Transiently Raised IOP Equivalent to That Experienced During Ocular Surgery Causes Moderate Inflammation but does not Affect Retinal Function or Result in Retinal Ganglion Cell Loss in An Animal Model
Abstract
Purpose:
High intraocular pressure (IOP) is known to result in retinal ganglion cell (RGC) loss, both with chronically raised intraocular pressure (such as with glaucoma) and with acute raises in pressure (due to injury or acute angle closure). Because IOP is often raised during ocular surgery, the purpose of this study was to evaluate the effect of transient moderate IOP on retinal function, RGC survival and the expression of Connexin 43 (Cx43) and glial fibrillary acidic protein (GFAP), ubiquitously expressed central nervous system (CNS) proteins that are known to be elevated during the retinal inflammatory response to injury.
Materials and Methods:
Wistar rats were exposed to transient IOP at 40 mmHg for 5 or 30 minutes, and 60 mmHg for 5 minutes (via cannulation of the anterior chamber with a saline reservoir raised to a height corresponding to the desired IOP), mimicking potential IOP rises during surgery such as DSAEK and some laser procedures (LASIK and femtosecond laser cataract surgery). Separate groups of animals had IOP maintained at 10 mmHg for 5 or 30 minutes as cannulation controls, or 120 mmHg for 60 minutes as positive controls. Changes in the optic nerve and retina were assessed immunohistochemically for GFAP and Cx43 expression. Retinal function was assessed using electroretinography (ERG) recorded at baseline and 14 days after the IOP rise and compared with RGC counts.
Results:
Results showed that there was a differential GFAP labelling pattern observed in the anterior optic nerve in the 40 mmHg 30 minute and 60 mmHg 5 minute groups 4 hours after manipulation. Gap junction protein Cx43 was minimally up-regulated in the retina in the short-term. There was, however, minimal long-term effect on retinal function and no RGC loss.
Conclusions:
n conclusion, elevations of IOP that are short in duration such as those occurring during surgical procedures, do not cause significant changes long-term in retinal function or RGC survival.
Key Messages:
Cx43 and GFAP are known to be elevated during the retinal inflammatory response to injury. No previous study has explored the effect of moderate and relatively short increases in IOP on the initial inflammatory response. We observed a mild glial inflammatory response in the anterior optic nerve, but only a minimal up-regulation of Cx43. However, transient and moderate IOP rises did not induce long term disruption to RGC function or number as measured by electrophysiology and RGC counts, respectively. This is applicable to clinical practice, as it means the IOP elevations that occur during some surgical procedures are unlikely to be causing long term damage in retinal function or RGC survival.
Article Information
- Received
- Accepted
- Published
Academic Editor: Yue Shi, University of California at Los Angeles
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2017 Jie Zhang, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Corresponding author: Helen V. Danesh-Meyer, Department of Ophthalmology, Faculty of Medical and Health Sciences, University of Auckland, Private bag 92019, Auckland 1142, New Zealand —
Competing Interests
The authors report no conflicts of interest.
Funding
No specific funding statement was provided by the authors.
Data Availability
No data-availability statement was provided by the authors.
Acknowledgements
This study was conducted with support from the Save Sight Society of New Zealand. The funding body had no involvement in the collection, analysis, and interpretation of data, in the writing of the report, or in the decision to submit the article for publication.
Citation:
Introduction
Transient and moderate increases in intraocular pressure (IOP) occur during many ophthalmic procedures. With Descemet’s stripping endothelial keratoplasty (DSEK) IOP increases of 30-40 mmHg are seen for 5-10 minutes,1, 2, 3. Other commonly performed ocular procedures produce shorter IOP rises, such as laser in-situ keratomileusis (LASIK) where the IOP increases to at least 65 mmHg for up to 45 seconds, and with femtosecond laser cataract surgery where increases in the IOP up to 40 mmHg for several minutes can occur,4.
Despite the known common occurrence of such IOP elevations, the effects on the optic nerve and retinal function have been minimally investigated. Recently, the role of glial behaviour with increases in IOP has been investigated and is suggested to be involved in retinal ganglion cell (RGC) dysfunction,5, 6. A variety of proinflammatory and inflammatory protein markers that are induced by IOP elevations have been identified,7. We have focused on the expression of the gap junction protein Connexin43 (Cx43) and the glial fibrillary acidic protein (GFAP) to evaluate the initial inflammatory response. GFAP is a marker for astrocytes and activated Müller cells. Astrocytes are recognised to have a role in controlling the microenvironment and providing structural and metabolic support to the optic nerve and retina,8, 9. Cx43 gap junction signalling and especially hemichannel opening plays a role in propagation of cell death signals after injury,10, 11, 12, 13, 14. Following injury to the CNS there is an astrocytic response that is well defined, where rapid proliferation is seen along with hypertrophy and changes in the expression of GFAP, as well as the upregulation of Cx43 expression,15, 16.
We have used electroretinogram (ERG) responses to measure retinal function. ERG responses in rats have been shown to be affected after IOP increase, with components of the ERG response showing differing levels of sensitivity to varying degrees of IOP elevation,17, 18. The negative and positive scotopic threshold response (nSTR and pSTR, reflecting retinal ganglion cell function) and photopic b-wave (bipolar cell function) were the most sensitive ERG components, being consistently reduced and delayed immediately after 75 minutes of IOP increase to 30 mmHg, and were completely abolished after 75 minutes of IOP at 70 mmHg,17. The most resilient ERG component was the a-wave (photoreceptor function), being the only component still present after 75 minutes of IOP at 70 mmHg,17. At IOPs above 70 mmHg, all components of the ERG were diminished,17. Therefore an IOP increase seems to affect cell function in the inner retina first, at an IOP level of 30 mmHg, followed by cell function in the middle and outer retina at progressively higher IOP levels.
Transient IOP increases above the systolic blood pressurefor more than 1 hour have been shown to result in retinal ganglion cell (RGC), inner retinal layer and outer nuclear layer cell apoptosis,19 and RGC loss has been reported in several other raised IOP models,20, 21, 22, 23, 24, 25, 26. The purpose of this study was to assess whether transient moderate elevated IOP relevant to those which patients are exposed to during ocular surgical procedures, modifies the optic nerve and retinal glial response, as well as RGC functional modification and RGC loss.
Materials and Methods
Animals
All procedures were conducted in compliance with the ARVO Statement on the Use of Animals in Ophthalmic and Vision Research and were approved by the Animal Ethics Committee of the University of Auckland. Ninety-six adult male Wistar rats weighing 250– 300 g were obtained from the Vernon Janson Unit at the University of Auckland and housed in a 12 hour light / 12 hour dark cycle and received food and water ad libitum. Light in the animal breeding unit was provided by white fluorescence light (Philips Master TLD 18W/965; Koninklijke Philips Electronics N.V., Shanghai, China). The light source covers broad band fluorescence, from 380 to 760 nm, and the average intensity at the top of the cage is 120 W/m2. Ninety animals had unilateral transient elevation of IOP to 10 mmHg for 5 or 30 minutes (cannulation controls, n = 10 per time point), 40 mmHg for 5 or 30 minutes (n = 20 per time point), 60 mmHg for 5 minutes, (n=20) and 120 mmHg for 60 minutes (n = 10). Uninjured animals (n = 6) were also used.
Transient Raised IOP Model
Animals were given an intraperitoneal injection of ketamine (60 mg/kg, Parnell Technologies, New Zealand) and medetomidine hydrochloride (0.4 mg/kg, Pfizer Animal Health, Australia) and the cornea anaesthetised with oxybuprocaine hydrochloride (0.4%, Bausch & Lomb, UK). The animal’s temperature was maintained using a heat pad. The left anterior chamber was cannulated with a 30-gauge infusion needle connected by silicone tubing to a reservoir of sterile 0.9% saline (Baxter, Australia). Cannulation was performed using a stereotaxic manipulator arm to avoid injury to the corneal endothelium, iris or lens. The saline reservoir was raised to a height corresponding to intraocular pressures of 10 mmHg for 5 or 30 minutes (normal IOP, cannulation control), 40 mmHg for 5 or 30 minutes, 60 mmHg for 5 minutes and 120 mmHg for 60 minutes (positive control based upon previous studies,26) . No saline leak from the eye was observed during the procedures corroborating that sustained pressure in the eye was achieved. IOP was measured using a TonoLab rebound tonometer (ICARE, Finland) 10 seconds after cannulation. The IOP obtained correlated well to that intended by calculation (a saline bag placed at a height of 13.595 cm produces an IOP of 10 mmHg, and so forth). The TonoLab tonometer has a measurement range of 1-99 mmHg, therefore inadequate for measurement of IOP at 120 mmHg. After the designated time period the cannula was removed. Experimental animals were either euthanised 4 hours later with CO2 and retinas cryosectioned for immunocytochemistry or were allowed to recover for 14 days for full-field ERG recording and RGC whole mount labelling.
Immunohistochemistry
Immunohistochemistry and confocal laser scanning microscopy (Olympus FV1000) were used to investigate the immediate effects of transient raised IOP on astrocytosis and Cx43 expression in the retina and anterior optic nerves 4 hours post IOP elevation. 4 hours after IOP elevation eyes were enucleated and fixed in 4% paraformaldehyde for 2 hours. The eyes were washed in PBS and transferred to sucrose prior to freezing in optimal cutting temperature compound (IA018, ProSciTech) using liquid nitrogen. For further details of methods refer to,26. Optic nerve containing sections (10 µm thick sagittal) were labelled for Cx43 using rabbit anti-Cx43 antibody (C6219, Sigma-Aldrich, 1:2000) and goat anti-rabbit IgG secondary antibodies conjugated to Alexa488 (A11034, Invitrogen; 1:500), and astrocytes were labelled using mouse anti-GFAP conjugated to CY3 (C9205, Sigma-Aldrich, 1:1000). Sections were incubated in the nuclear marker DAPI (40, 6-diamidino-2phenylindole, D9542, Sigma Aldrich, Germany) before being mounted using Citifluor AF-1 mounting medium (Citifluor, UK) and imaged.
GFAP intensity in the anterior optic nerve (the first 500 µm from the anterior end of the longitudinal section) as compared to the posterior optic nerve was assigned the category of either ‘strong’ or ‘normal’ by a masked observer. GFAP intensity in the GCL/NFL/OPL was also compared to posterior optic nerve intensity and assessed in the same way. To assess changes in Cx43, three locations were imaged from both the superior and inferior retina, starting one microscope field peripheral to the border of the optic disc, giving a total of six locations per retina. This method ensured similar locations were assessed between different eyes. At each location, two images were taken – one of the nerve fibre layer, ganglion cell layer and outer plexiform layer (NFL/GCL/OPL), and one of the inner nuclear layer, outer plexiform layer and outer nuclear layer (INL/OPL/ONL). The same settings were used within imaging of each retina. Owing to the low level of Cx43 in the normal retinas, imaging of the INL/OPL/ONL required a z-stack of 6 optical slices to be imaged, taken at 1 μm increments. Quantification of Cx43 was performed using automated spot counts in ImageJ software version 1.43 (National Institutes of Health, Bethesda, MD) as previously described,26.
GFAP intensity in the anterior optic nerve was analysed by binomial logistic regression using fluorescence (‘strong’ or ‘normal’) as the dependent variable and IOP and time of manipulation as covariates. A p-value less than 0.05 for each predictor indicates significance. A correlation test was also performed between fluorescence, IOP and time of manipulation. A p-value less than 0.05 for each pair tested indicated a significant correlation between the pair of variables.
Cx43 spot counts were analysed using multiple linear regression with average Cx43 spot count as the dependent variable and IOP and time of manipulation as independent variables. Student’s t-test was performed to further compare each experiment group to the uninjured group.
Retinal Ganglion Cell Counts
To investigate the effects of transient raised IOP on RGC survival in the long term, whole mount immunohistochemical techniques were used. For details refer to,25. Eyes were enucleated and the posterior eye cup was fixed in 4% paraformaldehyde in phosphate-buffered saline, (60 minutes, room temperature). The retina was then removed and permeabilised with 0.5% Triton X-100, (15 minutes, -80°C). Following thorough washing with PBS, free floating retinas were incubated overnight at 4°C in goat anti-Brn3a primary antibody (SC31984, Santa-Cruz Biotechnology, 1:100), 2% horse serum, 2% Triton X-100, PBS. After washing, a donkey anti-goat Cy3 (705-165147, Jackson Immuno Research, 1:500) secondary antibody solution was applied for 2 hours, room temperature. Nuclei were labelled with DAPI and retinas mounted onto SuperFrost-Plus slides (Thermo Scientific, US) using CitifluorTM mounting medium.
Imaging was with a confocal laser scanning microscope (Olympus FV1000). Two fields in each quadrant of each retina were imaged using a 10x objective lens giving a total of eight images per retina and total linear area of 6.5 mm2per retina. Quantification was performed using automated spot counts in NIH ImageJ software. RGC density was calculated as the number of RGCs per cm2. RGC density was compared to uninjured control and was analysed using Student’s t-test with a significance level of 0.05.
Electroretinograms
Full field ERGs were recorded from both eyes simultaneously at baseline and 14 days after IOP elevation. Animals were dark-adapted overnight and prepared for ERG recording under dim red lighting. Animals were anaesthetised with an intraperitoneal injection of ketamine (60 mg/kg) and medetomidine hydrochloride (0.4 mg/kg) and the cornea anaesthetised with oxybuprocaine hydrochloride (0.4%, Bausch & Lomb). The animal’s body temperature was maintained using a heat pad.
ERGs were recorded in a Faraday cage (custom-made) using a hand-held battery-operated ERG system (EPH-01, Ephios, Sweden) simultaneously from both eyes with silver / silver chloride electrodes (custom-made using a previously described method by Bui and Fortune) placed at the apex of the corneas, 27. The EPH-01 ERG system incorporates a signal amplifier, a filter set at 0.5-300 Hz, and an analogue to digital converter at 1 kHz, and records all data as waveforms for analysis by the included software MJOLNER. A ground platinum electrode was inserted into the base of the animal’s tail. Reference ring shaped silver / silver chloride electrodes (custom-made) were placed against the scleral conjunctiva behind the equator of both eyes. Eyes were lubricated using 0.5% carmellose sodium (Refresh Tears Plus, Allergan). Following electrode placement, Tropicamide eye drop (1%, Bausch & Lomb, UK) was instilled to induce mydriasis. Brief white LED flashes were delivered via a mini-Ganzfeld sphere integrated into the ERG unit and signal was recorded for five hundred ms after each flash by the ERG unit.
The scotopic threshold response (STR) was recorded at -5.1, -4.5, -4.2, and -3.9 log cd.s.m-2 with 10 flashes per light intensity with 2 second inter-stimulus intervals, and 10 second between-light intensity intervals,27, 28, 29. Intensities of -4.5, -4.2, and -3.9 log cd.s.m-2 produced consistent positive pSTR and negative nSTR. Scotopic ERGs at -3.05 and -2.7 log cd.s.m-2 was recorded with 10 flashes per light intensity with 2 second inter-stimulus intervals, and those at -2.2, -1.3, -0.4, 0.5, 1.4 log cd.s.m-2 were recorded as single flash responses, with 2 minute intervals between intensities and a 5 minute interval before the highest intensity achievable by the flash unit (2.0 log cd.s.m-2).ERG data were exported to the MJOLNER software supplied with the ERG unit. In the -5.1 to -2.7 log cd.s.m-2 range pSTR, b-wave, and nSTR amplitudes were measured from baseline to the peak or trough of the STR. Implicit time was estimated as the time required to reach the peak or trough of the STR. The STR response was used to evaluate RGC function as previously shown in a rodent glaucoma study,28. At intensities of -2.2, -1.3, -0.4, 0.5, 1.4, and 2 log cd.s.m-2, a-wave and b-wave amplitudes and implicit times were recorded from the automatic readings from the software. The software detects the a-wave amplitude and implicit time of the trough measured from baseline in the time window of 7-32 ms, and the b-wave amplitude and implicit time of the peak measured from the a-wave trough in the time window of 30-110 ms. The a-wave was used as an indication of photoreceptor function, and the b-wave ON bipolar cell function.
For each transient IOP treatment, 14 day ERG data at each flash intensity was compared to baseline from the same eyes of the same animals using paired sample two-tailed Student’s t-test. Two dependent variables (amplitude and implicit time) were tested against an independent variable - manipulation group. Because two outcome measures were tested against one hypothesised predictor, a Bonferroni-adjusted significance level of 0.025 was calculated to account for the increased probability of type one error. pSTR amplitude, pSTR implicit time, b-wave amplitude, b-wave implicit time, a-wave amplitude, and a-wave implicit time were compared.
Results
Expression of GFAP and Cx43 in the Optic Nerve And Retina
The optic nerve was labelled with GFAP and the pattern of labelling in each eye was analysed. A binomial logistic regression analysis of GFAP labelling intensity in the anterior (first 500 µm) and posterior optic nerve 4 hours after manipulation showed that both extent of IOP elevation (p=0.008, B=0.100, Exp(B)=1.105) and the duration (p=0.012, B=0.104, Exp(B)=1.110) were statistically significant predictors for a higher probability of strong GFAP labelling (Figure 1A-D). In accordance with this, the 120 mmHg 60 minute positive control group demonstrated the strongest GFAP fluorescence. Interestingly, strong GFAP fluorescence in the anterior optic nerve is seen at 10 mmHg (normal IOP) for 30 minutes, when none is seen at 40 mmHg for 5 minutes (Figure 1E). This would suggest that the cannulation is having some effect on the retina and indicates that the duration of the manipulation is an important factor in producing change. Analysis of GFAP intensity in the NFL/GCL/OPL in the retina was carried out in the same way and no statistically significant difference was found. Analysis of Cx43 spot count (number of spots per 210 µm linear image) in the NFL/GCL/IPL showed that the degree of IOP elevation was a statistically significant predictor of Cx43 spot count (p<0.05, B=0.344) in a multiple linear regression analysis, whereas duration of IOP elevation (p=0.44) was not (Figure 2). However, no significant difference was found between any of the moderate IOP manipulation groups and uninjured retinas using a Student’s t-test. A significantly higher Cx43 spot count was found in the 120 mmHg 60 minute group positive control compared to uninjured retinas (p<0.050, 79 ± 9.7 vs. 42 ± 10) using a Student’s t-test. The same analysis of Cx43 spot count in the INL/OPL/ONL did not reveal significant differences. No significant differences were found in Cx43 spot count in the optic nerve (Supplementary figure 1).
Figure 1. GFAP labelling (red) in the optic nerve 4 hours post IOP manipulation. Blue labelling is DAPI for cell nuclei. (A) An optic nerve from the uninjured (i.e. completely naïve) group showing uniform GFAP fluorescence in both the anterior (first 500 µm) and posterior optic nerve. (B) An optic nerve from the 120 mmHg 60 minute group showing much stronger GFAP labelling in the anterior part of the optic nerve compared to the posterior part - this sample was assigned to the ‘strong’ labelling group. (C) Optic nerves from the 40 mmHg 30 minute group and (D) 60 mmHg 5 minute group showing stronger GFAP in the anterior optic nerve compared to the posterior optic nerve. Scale bar = 500 µm. (E) Graph showing the percentage of animals with normal (pale grey) or strong (dark grey) GFAP fluorescence in the anterior optic nerve (first 500 µm) examined 4 hours after transiently raised IOP for different periods of time. n = 6, 5, 5, 10, 10, 10, 5, respectively, from left to right.
Download figure
Figure 2. Representative confocal microscope single optical slice images of retina sagittal sections labelled for Cx43 (green), GFAP (red), and DAPI for nuclei (blue) and graphed Cx43 spot counts. (A) An uninjured retina showing regular glial processes labelled with GFAP in the NFL and Cx43 dots with some co-localisation in the same layer (resulting in yellow label). (B) A retina 14 days after 120 mmHg 60 minute manipulation shows reactive glial processes (red) and an increased amount of Cx43 label. (C) Retinas 14 days after 40 mmHg 30 minute and (D) 60 mmHg 5 minute manipulations did not change the number of Cx43 immunoreactive labelling spots or cause astrocytosis in the retina after 14 days. Scale bar = 100 µm. (E) Graph showing Cx43 spot counts in the NFL/GCL/IPL. n=6, 5, 5, 10, 10, 10, 5 for each group from left to right.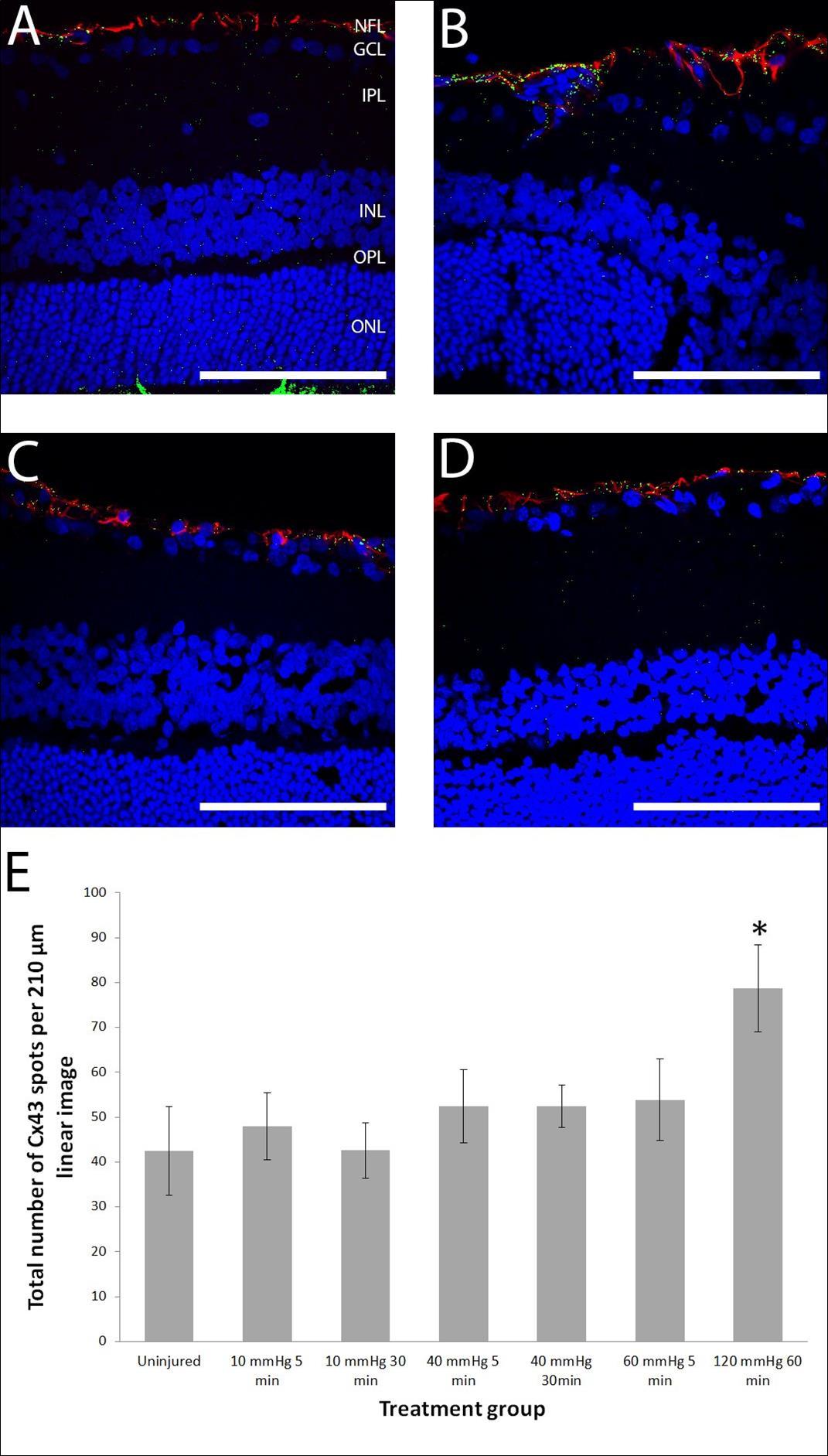
Download figure
Retinal Ganglion Cell Density
The transient periods of raised IOP at 40 and 60 mmHg did not affect RGC density at the end of 14 days (Figure 3). A significant decrease (p<0.001) in RGC density was found in the positive control group of animals where the IOP was raised to 120 mmHg for 60 minutes (115632 ± 6173 cells/cm2) compared to uninjured (218771 ± 4582 cells/cm2), 10 mmHg (normal IOP cannulation) for 5 minutes (228308 ± 26611 cells/cm2, or 10 mmHg for 30 minutes (normal IOP cannulation) (232035 ± 7970 cells/cm2). No significant decrease in other treated groups was found compared to uninjured, 10 mmHg for 5 minutes or 10 mmHg for 30 minutes.
Figure 3. Graphed results displaying RGC density 14 days post raised intraocular pressure and confocal microscope single optical slice images of flat mounted retinas with Brn3a labelled RGCs. (A) Average RGC density 14 days after transient raised intraocular pressure. *A significant decrease was found in the group subjected to 120 mmHg intraocular pressure for 60 minutes compared to the uninjured group. n = 5, 3, 5, 10, 8, 10, 5 for each group from left to right. Images shown (B-E) were taken from the superior periphery of the retina (B) An uninjured retina is seen with densely packed RGCs. (C) A retina 14 days after 120 mmHg 60 minute manipulation had a reduced RGC density. (D) 40 mmHg 30 minute and (E) 60 mmHg 5 minute manipulations did not reduce RGC density. Scale bar = 500 µm.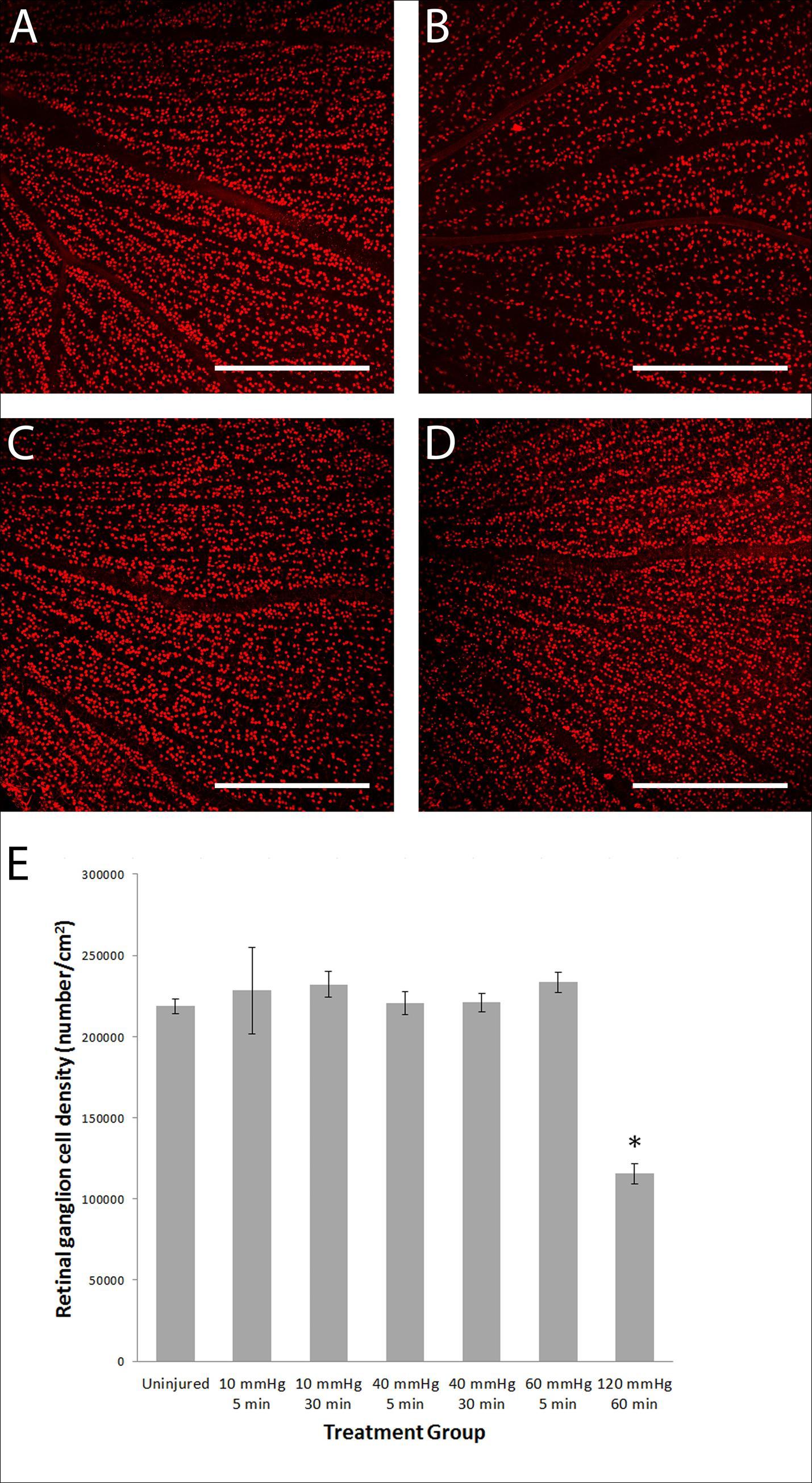
Download figure
Effect of Transient Intraocular Pressure Rise on Retinal Function
Retinal functional modifications were assessed 14 days after transiently raised IOP (Figure 4). Figure 4A gives examples of graphs that show how measurements were read from an ERG recording and Figure 4B shows examples of ERG waveforms in the scotopic threshold range and the b-wave range. There was very limited evidence for a decrease in pSTR (-5.1 to -3.9 log cd.s.m-2) and b-wave (-3.05 to 2.0 log cd.s.m-2) amplitude with cannulation and after transient IOP rise to 40 and 60 mmHg regardless of the duration of the IOP rise (Figure 4C and D). A smaller b-wave amplitude with significant difference compared to the baseline was only found in the 120 mmHg 60 minute positive control group at -2.7, -2.2, -1.3, and 1.4 log cd.s.m-2, including 2 animals out of 4 that completely lost their pSTR and b-wave response from -5.1 to -2.7 log cd.s.m-2. pSTR amplitude (Supplementary figure 2) and b-wave amplitude (Supplementary figure 3) of the contralateral eyes at 14 days were compared to their respective baselines (the same eyes recorded before manipulation), and no significant difference was found in any group.
No significant change in pSTR or b-wave implicit time was found in the manipulated eyes of any group. There was also no significant change in nSTR, a-wave amplitude (Supplementary figure 4) or implicit time for any group compared to baseline.
Figure 4. Full-field ERG findings 14 days after a transient IOP increase. (A) Graphs showing how the various measurements were read from an ERG recording. pSTR amplitude and implicit time were measured from the baseline (0 µV and 0 ms) to the peak. nSTR amplitude and implicit time were measured from the baseline to the trough. a-wave amplitude and implicit time were measured from the baseline to the trough. b-wave amplitude was measured from the trough of the a-wave to the peak of the b-wave. b-wave implicit time was measured from the baseline to the peak. (B) Examples of ERG waveforms in the scotopic threshold range (first 4 rows) and the b-wave range (last 8 rows). pSTR amplitudes (C), and b-wave amplitude (D) measured 14 days after transiently raised IOP for different durations. Data from all cannulated eyes recorded before cannulation was pooled to give a representation of baseline pSTR or b-wave amplitudes at different flash intensities (black lines). * indicates significant differences compared to baseline at the particular flash intensity at a significance level of 0.025. (C-D) 10 mmHg 5 min n=5, 10 mmHg 30 min n=5, 40 mmHg 5 min n=10, 40 mmHg 30 min n=10, 60 mmHg 5 min n=9, 120 mmHg 60 min n=4, Baseline n=43. Error bars represent SEM.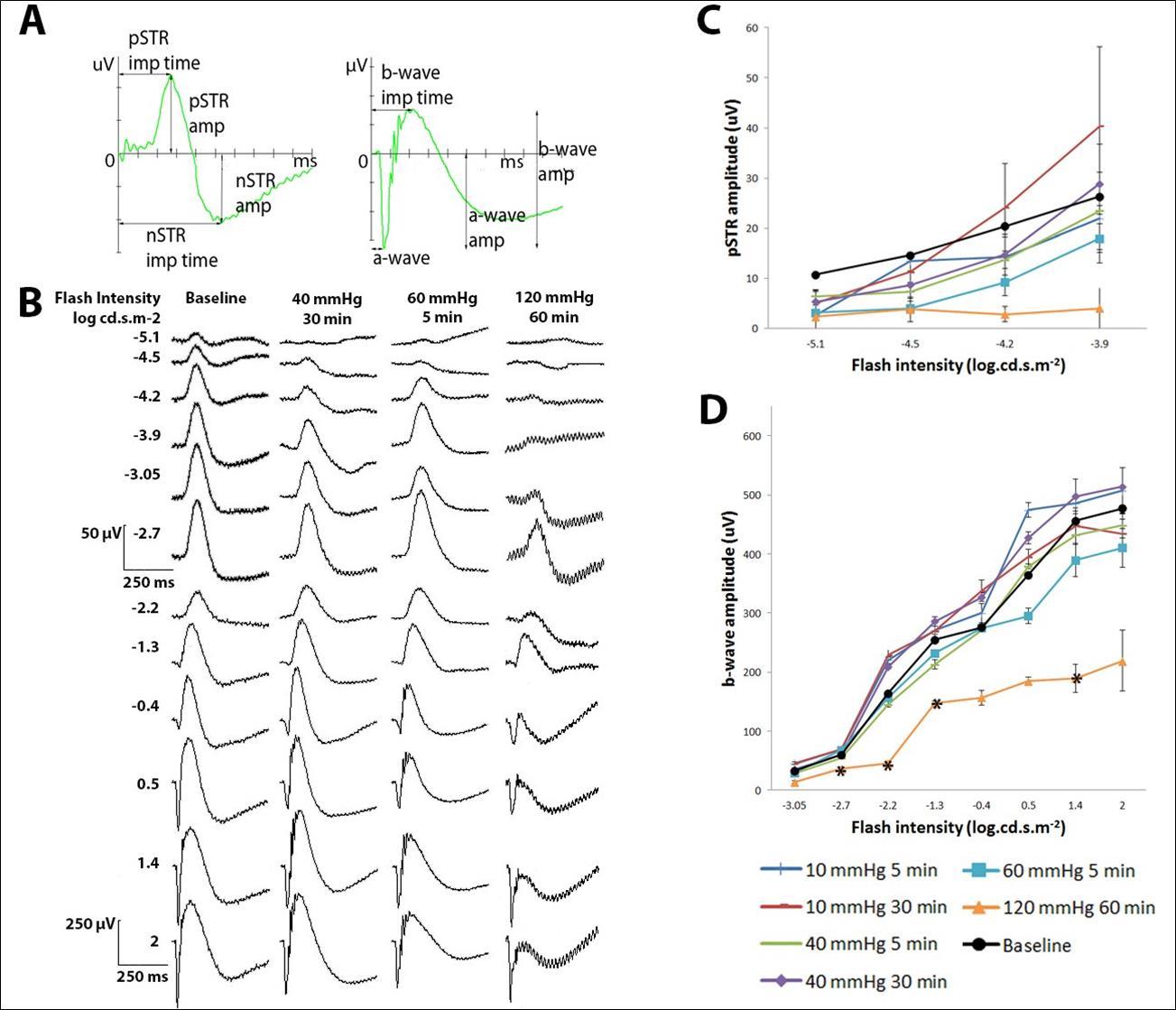
Download figure
Discussion
This study suggests that transient and moderate increases in IOP (40-60 mmHg over several minutes), similar to those that occur during many ophthalmic procedures such as DSEK, LASIK and femtosecond laser cataract surgery,1, 2, 3, 4, do not result in permanent functional retinal damage. This study extends the work of other investigators studying the effect of acute IOP elevations on retinal ERG responses. We did not see any change in ERG responses at 14 days, except in the positive control where IOP was raised to 120 mmHg for 60 minutes. Other investigators have considered the 30 minute time point or longer and have not seen permanent damage with moderate increases in IOP. In one study the pSTR significantly changed one week after elevation of IOP to 50 mmHg for 30 minutes, but the b-wave completely recovered,30. With higher IOP (50-70mmHg for 15-60 minutes) greater loss and slower recovery in the nSTR response,31 has been demonstrated. One further study with an 8 hour, single episode of moderate IOP elevation (50 mmHg) reported no disruption to either anterograde or retrograde axonal transport at any time point beyond the acute injury event, or long term damage to RGCs,32. In general there seems to be a threshold for permanent RGC functional loss in the rat at an IOP between 60-70 mmHg if sustained for 105 minutes or more,33, or with shorter time periods at higher pressures (120 mmHg sustained for 60 minutes),25.
No previous study has explored the effect of moderate and relatively short increases in IOP on the initial inflammatory response. We observed a mild glial inflammatory response in the anterior optic nerve. Astrocytosis was identified in the anterior optic nerve in the 40 mmHg 30 minute and 60 mmHg 5 minute groups 4 hours after manipulation, but not in the 40 mmHg 5 minute group. Both the degree and duration of the IOP elevation were significant predictors of astrocytosis, and functional changes in RGCs were seen when the IOP was raised to higher levels in the positive control group (120mmHg for 60 minutes). However, as in the Abbott et al. study, transient and moderate IOP rises did not induce long term disruption to RGC function or number as measured by electrophysiology and RGC counts, respectively.
There is increasing evidence that glial cells play a pivotal role in modulating the microenvironment of the optic nerve. GFAP is recognised to be a sensitive marker of astrocyte activation in response to injury,34 with activation of glial cells in the optic nerve head in experimental glaucoma models as well as in humans with glaucoma, 35, 36, 37, 38. Previously, we have shown that ischemic IOP levels (120mmg, 60 minutes) led to activation of astrocytes and Müller cells in the retina at 8 hours and 3 days post-injury,25, 39. In that study, as in the present study positive control group, this is associated with subsequent RGC death.
Cx43 has been shown to be up-regulated in both the GCL and anterior optic nerve in human glaucoma retinas,37 and in the GCL 3 days after animals were subjected to partial optic nerve transection, coinciding wit with the timing of RGC loss,40. An ischaemic IOP increase to 120 mmHg for 60 minutes resulted in Cx43 up-regulation in the GCL from 4 hours, co-localising with activation of glial cells and preceding RGC loss at 21 days,25. The range of transient moderate IOP increase tested in the current study appeared insufficient to cause Cx43 up-regulation in the GCL and anterior optic nerve at 4 hours, and was not associated with any RGC loss at 14 days. The threshold of IOP elevation and/or ischemia necessary to induce retinal Cx43 and GFAP changes needs further evaluation.
It is of note that cannulation alone can cause an inflammatory response,41. In that study cannulation of the anterior chamber alone resulted in increased hyalocyte density, microglical activation and accumulation of macrophages in the subretinal space. These increases were also seen after IOP elevation (50 mmHg for 30 minutes) with ERG signals (derived from RGCs) reduced significantly in those eyes experiencing pressure elevation, but not cannulation only. In this present study there was an increase in GFAP labelling in the anterior optic nerve head with sham cannulation (10 mmHg (normal IOP) for 30 minutes) and like the above study we did not observe cannulation effects on ERG recordings.
To make comparisons between the human and rat eye, one should consider the differences in anatomy and morphology. In the rat, the diameter of the optic nerve head is smaller than in humans (about nine times smaller),42. Like humans, the rat does have a lamina cribrosa-like structure, but the quantity of the lamina cribrosa seems to be dependent on rat strain. Collagen types I, III, and IV are found in the lamina cribrosa. In rats the lamina cribrosa is composed of more type IV collagen fibres than humans. Type I and III fibres are interstitial collagens, whereas type IV is a basement collagen,43. Thus, there may be differences in the biomechanical properties of the lamina cribrosa between these species. However, a study done by one group looked at the changes in the optic nerve head in aged rats and found that the changes were similar to those found in the human lamina cribrosa, thus concluding that the rat model was useful for studying the influence of age on the onset of glaucoma,42. The blood vessels (central retinal artery and vein) in rats enter the optic nerve head at the level of the sclera as opposed to retrolaminar in humans, predisposing the retinal blood supply to IOP rises more than in humans. The blood supply of the optic nerve in rats is via the central retinal artery and possibly the pia vessels, whereas in humans the blood supply is via the central retinal artery, pia vessels, and possibly via the choroid. These suggest that rat retinas may be more prone to the effects of transient IOP increases than human retinas, even though more experimental evidence is needed. Lastly the ganglion cell layer (GCL) morphology differs between humans and rats, with the average cell number being between 72,000 and 113,000 in rats and between 700,000 and 1,500,000 in humans,42.
In humans ophthalmic surgery is usually carried out under local anaesthetic, so the impact of general anaesthesia on RGC survival should also be taken into consideration. A number of studies have looked at the effect of general anaesthesia on retinal response and it has been found that there are differing effects on the ERG depending on the anaesthetic used. One group showed that ketamine/xyalazine with pancuronium was the best combination to minimise eye movement and maximise retinal function,44. While another group showed that Medetomidine prolonged the implicit time and lowered the amplitude response of the a- and b-wave at all intensities, but clinically only minimally affected the retinal responses,45. It has also been shown that pre-treatment with ketamine/xylazine protected the retina against cell damage, reducing photoreceptor death, suggesting that anaesthesia may provide neuroprotective effects in light induced cell damage,46. With advances in technology it may be possible, in the future, to perform ERG studies in animals without requiring general anaesthesia and thus removing the confounding effect of it.
Our laboratory is particularly interested in the role of connexins and astrocytic regulation after injury due to intraocular pressure increases, hence the focus on this marker of inflammation. However, future directions may also involve the examination of various other genes (and their proteins), such as the pro-inflammatory and inflammatory genes Bcl2, Birc4/XIAP, Cat and SAA1 genes, whose transcription levels have been shown to be altered by elevations in IOP,7.
A limitation of the study is the use of a semi-quantitative grading system, even though a masked observer analysed the images collected in order to reduce subjectivity. Other molecular biology approaches including PCR and Western Blotting may be useful in studying connexin43 and GFAP gene and protein expression changes.
Future studies should aim to more clearly define the threshold of IOP elevation and/or ischemia that will induce permanent retinal changes and subsequent permanent vision loss. This should include aged animals with IOP levels and durations pertinent to cataract surgery (40-80 mmHg for 15-30 minutes). There have been reports of increased RGC susceptibility to permanent injury after IOP elevation and sham cannulation insult in older rat eyes,47.
In conclusion, the range of transient moderate IOP changes in ophthalmic procedures such DSEK and LASIK may provoke a glial response in the optic nerve, but no significant changes in Cx43 expression. We have yet to define how persistent this response might be. No significant retinal functional changes or reduction in RGC number was found. These surgical procedures therefore appear to be relatively safe.
Supplementary Figures:
References
- 1.Karimian F, Sadoughi M M. (2010) Air‐assisted descemet stripping automated endothelial keratoplasty with posterior chamber fixation of an aphakic iris‐claw lens. , J Ophthalmic Vis Res; 5, 205-10.
- 2.Liang C M, Chen Y H, Lu D W.New continuous air pumping technique to improve clinical outcomes of descemet‐stripping automated endothelial keratoplasty in asian patients with previous ahmed glaucoma valve implantation. PLoS One 2013;8:e72089.
- 3.Price F W, Price M O. (2006) Descemet’s stripping with endothelial keratoplasty in 200 eyes: early challenges and techniques to enhance donor adherence. , J Cataract Refract Surg; 32, 411-18.
- 4.Kerr N M, Abell R G, Vote B J. (2013) Intraocular pressure during femtosecond laser pretreatment of cataract. , J Cataract Refract Surg; 39, 339-42.
- 5.Burgoyne C F. (2011) A biomechanical paradigm for axonal insult within the optic nerve head. , Exp Eye Res; 93, 120-32.
- 6.Mandal A, Shahidullah M, Delamere N A. (2007) Elevated hydrostatic pressure activates sodium/hydrogen exchanger‐1 in rat optic nerve head astrocytes. , Am J Physiol Cell Physiol; 297, 111-20.
- 7.Walsh M M, Yi H, Friedman J. (2009) Gene and protein expression pilot profiling and biomarkers in an experimental mouse model of hypertensive glaucoma. , Exp Biol Med; 234(8), 918-30.
- 8.Lye-Barthel M, Sun D, Jakobs T C. (2013) Morphology of Astrocytes in a Glaucomatous Optic Nerve. Invest Ophthalmol Vis Sci2013;. 54, 909-17.
- 9.Nedergaard M, Ransom B, Goldman S A. (2003) New roles for astrocytes: redefining the functional architecture of the brain. , Trends Neurosci2003; 26, 523-30.
- 10.O’Carroll S J, Gorrie C A, Velamoor S.Connexin43 mimetic peptide is neuroprotective and improves function following spinal cord injury.NeurosciRes2013;75(3):. 256-67.
- 11.Lin J H, Weigel H, Cotrina M L.Gap-junction-mediated propagation and amplification of cell injury.NatNeurosci1998;1:. 494-500.
- 12.Danesh-Meyer H V, Zhang J, Acosta M L.. Connexin43 in retinal injury and disease. Prog Retin Eye Res2015October9 .
- 13.Cronin M, Anderson P N, Cook J E.Blocking connexin43 expression reduces inflammation and improves functional recovery after spinal cord injury.MolCellNeurosci2008;39:. 152-60.
- 14.Rawanduzy A, Hansen A, Hansen T W.Effective reduction of infarct volume by gap junction blockade in a rodent model of stroke.JNeurosurg2009;87:. 916-20.
- 15.Rouach N, Avignone E, Meme W.Gap junctions and connexin expression in the normal and pathological central nervous system.BiolCell2002;94:. 457-75.
- 16.Lee I H, Eva L, Ole K.Glial and neuronal connexin expression patterns in the rat spinal cord during development and following injury.JCompNeurol2005;489:. 1-10.
- 17.Bui B V, Edmunds B, Cioffi G A.The gradient of retinal functional changes during acute intraocular pressure elevation. InvestOphthalmolVisSci2005;46: 202-13.
- 18.Chrysostomou V, Crowston J G.The photoic negative response of the mouse electroretinogram: reduction by acute elevation of intraocular pressure. Invest Ophthalmol Vis Sci 2013 July;54(7): 4691-97.
- 19.Ju W K, Kim K Y.Measuring glutamate receptor activation‐induced apoptotic cell death in ischemic rat retina using the TUNEL assay.MethodsMolBiol2011;740:. 149-56.
- 20.Hanninen V A, Pantcheva M B, Freeman E E.Activation of caspase 9 in a rat model of experimental glaucoma.CurrEyeRes2002;25:. 389-95.
- 21.Jha P, Banda H, Tytarenko R.Complement mediated apoptosis leads to the loss of retinal ganglion cells in animal model of glaucoma.MolImmunol2011;48:. 2151-58.
- 22.Lam T T, Kwong J M, Tso M O.Early glial responses after acute elevated intraocular pressure in rats. InvestOphthalmolVisSci2003;44: 638-45.
- 23.McKinnon S J, Lehman D M, Kerrigan-Baumrind L A.Caspase activation and amyloid precursor protein cleavage in rat ocular hypertension. , Invest Ophthalmol Vis Sci 2002, 1077-87.
- 24.Sawada A, Neufeld A H.Confirmation of the rat model of chronic, moderately elevated intraocular pressure. ExpEyeRes1999;69: 525-31.
- 25.Danesh-Meyer H V, Kerr N M, Zhang J.Connexin43 mimetic peptide reduces vascular leak and retinal ganglion cell death following retinal ischaemia.Brain2012;135:. 506-20.
- 26.Kerr N M, Johnson C S, Zhang J.High pressure induced retinal ischaemia reperfusion causes upregulation of gap junction protein connexin43 prior to retinal ganglion cell loss.ExpNeurol2012;234:. 144-52.
- 27.Bui B V, Fortune B.Ganglion cell contributions to the rat full‐field electroretinogram.JPhysiol2004;555:. 153-13.
- 28.Fortune B, Bui B V, Morrison J C.Selective ganglion cell functional loss in rats with experimental glaucoma. InvestOphthalmolVisSci2004;45: 1854-62.
- 29.Saszik S M, Robson J G, Frishman L J.The scotopic threshold response of the dark‐adapted electroretinogram of the mouse.JPhysiol2002;543:. 899-916.
- 30.Kong Y X, Crowston J G, Vingrys A J. (2009) Functional changes in the retina during and after acute intraocular pressure elevation in mice.InvestOphthalmolVisSci2009;50:. 5732-40.
- 31.He Z, Bui B V, Vingrys A J.The rate of functional recovery from acute IOP. elevation.InvestOphthalmolVisSci2006;47: 4872-80.
- 32.Abbott C J, Choe T E, Lusardi T A.Evaluation of retinal nerve fiber layer thickness and axonal transport 1 and 2 weeks after 8 hours of acute intraocular pressure elevation in rats.InvestOphthalmolVisSci2014;55:. 674-87.
- 33.Bui B V, Batcha A H, Fletcher E.Relationship between the magnitude of intraocular pressure during an episode of acute elevation and retinal damage four weeks later in rats. PLoS One 2013;8:e70513.
- 35.Balaratnasingam C, Morgan W H, Bass L.Elevated pressure induced astrocyte damage in the optic nerve.BrainRes2008;1244:. 142-54.
- 36.Furuyoshi N, Furuyoshi M, May C A.Vascular and glial changes in the retrolaminar optic nerve in glaucomatous monkey eyes.Ophthalmologica2000;214:. 24-32.
- 37.Kerr N M, Johnson C S, Green C R.Gap junction protein connexin43 (GJA1) in the human glaucomatous optic nerve head and retina.JClinNeurosci2011;18:. 102-08.
- 38.Varela H J, Hernandez M R.Astrocyte responses in human optic nerve head with primary open‐angle glaucoma.JGlaucoma1997;6:. 303-13.
- 39.Zhang S D, Li W Y, Wang W Q.Detection of early glial responses in the retina, optic nerve and optic chiasm in acute ocular hypertension rat model.ChinJExpOphthalmol2012;30:. 704-08.
- 40.Chew S S, Johnson C S, Green C R.. Response of retinal Connexin43 to optic nerve injury.InvestOphthalmolVisSci2011;52: 3620-29.
- 41.Kezic J M, Chrysostomous V, Trounce I A.Effect of anterior chamber cannulation and acute IOP elevation on retinal macrophages in the adult mouse.InvestOphthalmolVisSci2013;54:. 3028-36.
- 42.Albrecht May C.Comparative anatomy of the optic nerve head and inner retina in non-primate animal models used for glaucoma research.OpenOphthalmolJ2008;2:. 94-101.
- 43.Rehnberg M, Ammitzböll T, Tengroth B.Collagen distribution in the lamina cribrosa and trabecular meshwork of the humaneye.BrJOphthalmol1987;71(12):. 886-92.
- 44.Nair G, Kim M, Nagaoka T.Effects of common anaesthetics on eye movement and electroretinogram.DocOphthalmol2011;122(3):. 163-76.
- 45.Norman J C, Narfström K, Barrett P M.The effects of medetomidine hydrochloride on the electroretinogram of normal dogs.VetOphthalmol2008;11(5):. 299-305.
Cited by (4)
This article has been cited by 4 scholarly works according to:
Citing Articles:
Rachel S. Chong, J. M. Busoy, Bingyao Tan, S. Yeo, Ying Shi Lee et al. - Translational Vision Science & Technology (2020) Semantic Scholar OpenAlex
Investigative Opthalmology & Visual Science (2019) Crossref OpenAlex
Yamunadevi Lakshmanan, F. S. Wong, Wing-yan Yu, S. Li, K. Choi et al. - Investigative Ophthalmology and Visual Science (2019) Semantic Scholar