Adaptive Contribution of Thyroid Hormones in Obesity
Abstract
Nowadays, obesity is seriously threatening healthy life in all societies, especially in developed societies. Obesity is showing its likely effects through biologically active substances called adipokines. The most familiar of these is leptin. Leptin is synthesized in directly proportional to the amount of adipose tissue and reduced appetite by stimulating the satiety center. This status is like a protective mechanism that tries to reverse severe pathological process. Similar to this behavior of leptin, thyroid hormones are increasing in the advanced stages of obesity, increasing the resting energy expenditure (REE). The accelerating oxidative phosphorylation causes the use of energy as heat, the energy that has not transformed into ATP, together with ATP synthesis. This situation, the significant portion of energy provides to consumed instead of storing as fat. In addition, finding that T3 accelerates glucose transport and the TCA cycle without changing the rate of ATP synthesis in skeletal muscles suggests that thyroid hormones may be an effective tool in standing against obesity. In addition of that, the presence of studies indicating that thyroid hormones have an increasing tendency in the advanced stages of obesity is likely thought to be a rescuer mechanism to increase the effectiveness of suppressed thyroid hormones. On the contrary of these ideas, having been reported suppressing 5’-deiodinease enzyme activity in chronic diseases causes anxiety about the effectiveness of thyroid hormones in obesity. Based on available information, we aimed to prepare a review evaluating of this adaptive condition of thyroid hormones.
Article Information
- Received
- Accepted
- Published
Academic Editor: Kingsley Appiah, Kumasi Technical University, Ghana.
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2019 Fatih Ozcelik, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Corresponding author: Fatih Ozcelik, Department of Medical Biochemistry, University of Health Sciences, Faculty of Medicine, 34668, Istanbul, Turkey —
Competing Interests
The authors have declared that no competing interests exist.
Funding
No specific funding statement was provided by the authors.
Data Availability
No data-availability statement was provided by the authors.
Citation:
Introduction
Nowadays, obesity has become the leading health problem with its increasing prevalence day by day. Because of the effects on all systems in the advanced stages of obesity is seriously threatening healthy life. In addition, thereby obesity cause significant effects on endocrine, cardiovascular, respiratory, gastrointestinal and leukomotor systems, increasing the morbidity and mortality of the society. With a simple definition is that obesity is an observed situation a result of taking more energy than needed. Genetic structure, endocrine conditions affecting the metabolism (such as Cushing's disease, hypothyroidism and polycystic ovary syndrome), nutritional habits, the psycho-social and cultural factors the interaction of one or more these factors are the main causes of obesity. Obesity, which is characterized by an abnormal increase in the amount of adipose tissue, is the most common chronic disease of our age in developed and developing countries 1, 2. According to the latest data shared by the World Health Organization (WHO), the incidence of obesity is increasing among European countries as in the USA. It was found that obesity, having become a serious public health problem in developed societies was associated with insulin resistance, diabetes, atherosclerosis, hypertension, chronic kidney disease, and increasing cardiovascular morbidity and mortality 3, 4.
The main role of adipose tissue is to store energy. It fulfills this role by storing fat transported through lipoproteins or by synthesizing fatty acid from glucose 5. In addition, physical preservation, as well as the storage of fat-soluble vitamins (vitamin A, D, E and K) and providing body temperature hemostasis are the other roles of adipose tissue.
Synthesis and Effects of Adipokines in Adipocytes
Adipose tissues are composed from adipocytes, fibroblasts, leukocytes and macrophage cells. There are two types of adipose tissue in the body: white adipose tissue and brown adipose tissue. The fact that leptin from adipokines having been found first time in 1994 showed that white adipose tissue was also an endocrine organ 6. Since then, white adipose tissue has been found to release many adipokines having several functions. Interleukin 6 (IL-6) and tumor necrosis factor alpha (TNF-α) secreted from macrophages in adipose tissue were also counted among adipokines. IL-6 has been found to have a regulating effect on other adipokines and reported that decrease adiponectin levels 7. Likewise, TNF-α has been found to have regulatory effects on other adipokines (reducing adiponectin levels and increasing leptin levels). With reduced the development of obesity, and decreased insulin and leptin levels have been reported in animal studies in which the effect of TNF-α was blocked 8, 9.
Recently, adipokines have been found to be secreted from other cells, except for adipocyte cells. Furthermore, adipokines associated with adipose tissue were indicated to have an important role in progression of insulin resistance, hypertension, inflammation, cardiovascular diseases. The recent year studies have been confirmed that white adipose tissue is an active endocrine organ that secretes adipokines, which are bioactive polypeptides. The complexity of the effects of adipokines on the body caused many researchers to have different or even contradictory results. It has been reported that adipokines affect insulin sensitivity, oxidative capacity and lipid intake in the peripheral, while regulating appetite and energy expenditure in the central 10, 11, 12, 13, 14, 15. After the emergence that adipose tissue produces/secretes a group of bioactive polypeptides, which are defined as adipokines, studies on the negative effects of obesity through these mediators have accelerated. However, on the contrary of other, the presence of adipokines, such as leptin (with its inhibitory effect against weight gain) than other adipokines, has brought a different dimension to the discussions. Thus, leptin is mainly produced from adipocytes in subcutaneous white adipose tissue and it has been thought to control the growth of adipose tissue by affecting the satiety center in the central nervous system 16, 17, 18.
Studies in animals with gene mutation responsible for leptin production have shown that obesity develops due to gene mutation, and again weight gain is stopped and even weight loss occurs when leptin is administered to these animals. These findings suggested that leptin may be an important mediator in controlling obesity. In another recent study, founding high levels of blood leptin in a significant proportion of obese patients suggested that there might be a rescuer endocrine response developed by the body's adipose tissue against obesity. In this context, the presence of obese patients with increased weight despite hyperleptinemia suggests that resistance to endogenous leptin may develop. In studies supporting this idea, have been found that some proteins increase with causing resistance against to endogenous leptin and inhibiting leptin-dependent stimulation. These mechanisms are indicated by the fact that leptin cannot act on the target neurons and does not cross the blood-brain barrier (BBB). A clinical-based study reported that leptin contributed to the regulation of blood glucose levels and increased insulin sensitivity 13, 19. In addition it has also been shown that leptin is associated with the formation of TNF-α and reactive oxygen products, macrophage activation and synthesis of nitric oxide synthase. These findings 20, 21, 22, also shows that obesity-related hyperleptinemia may have a harmful effect on the cardiovascular system.
In a study investigating the association of adipose tissue amount and location with circulating adiponectin levels, it was shown that adiponectin concentration decreased with obesity and increasing amount of visceral adipose tissue, but increased adiponectin concentration with weight loss 23. In addition, adipose tissue may also release different mediators or markers than adipokine that may be associated with obesity. Ghrelin, serum amyloid A, resistin, retinol binding protein 4 (RBP-4), vaspin, visfatin, omentin and chemerin are among these mediators. An example is the fact that plasminogen activator inhibitor-1 (PAI-1), which is primarily secreted from hepatocytes and endothelium, has been reported to be secreted from visceral white adipose tissue. In a study of patients have central obesity have found that the high level of PAI-1 concentration and the PAI-1 concentrations decrease in weight loss 24, 25, also support this finding. In a different study on PAI-1 deficient mice, obesity and insulin resistance were not observed. This condition supports the relationship between PAI-1 levels and adipose tissue 26. In a multicenter insulin resistance-atherosclerosis study involving different ethnic groups aimed at investigating the relationships between insulin resistance, cardiovascular risk factors and disease, a correlation was found between increased plasminogen activator inhibitor-1 levels and the development of type 2 diabetes 27. All these findings suggest that people with high amount of adipose tissue are prone to thrombosis (thrombophilia). This information explains why obesity has a high mortality risk.
Energy Consumption in Obesity and the Relationship with Thyroid Hormone
It is known that TRH, TSH and T3 are actively involved in the regulation of the release of thyroid hormones (Figure 1). The hypothalamic-pituitary-thyroid (HPT) axis has also been shown to be regulated by adipose tissue-dependent leptin and nutritional status 18, 28, 29. In such a regulation, upon considering the regulatory effect of the thyroid on thermogenesis, it has been suggested that thyroid hormones might be effective in preventing of the development of obesity 18, 30. It is known that TSH is significant on energy metabolism, especially in 3,5-diiodo-l-thyronine (3,5-T2), T3 and T4 levels 30, 31. However, conclusive evidences about its obesity-inhibiting contributions are needed.
Figure 1. Thyroid hormones synthesis. The synthesis of thyroid hormones begins with hypothalamic stimulation. Increased level of T4 or T3 hormones suppress basal TSH levels by negative feedback and inhibit the TSH response to TRH.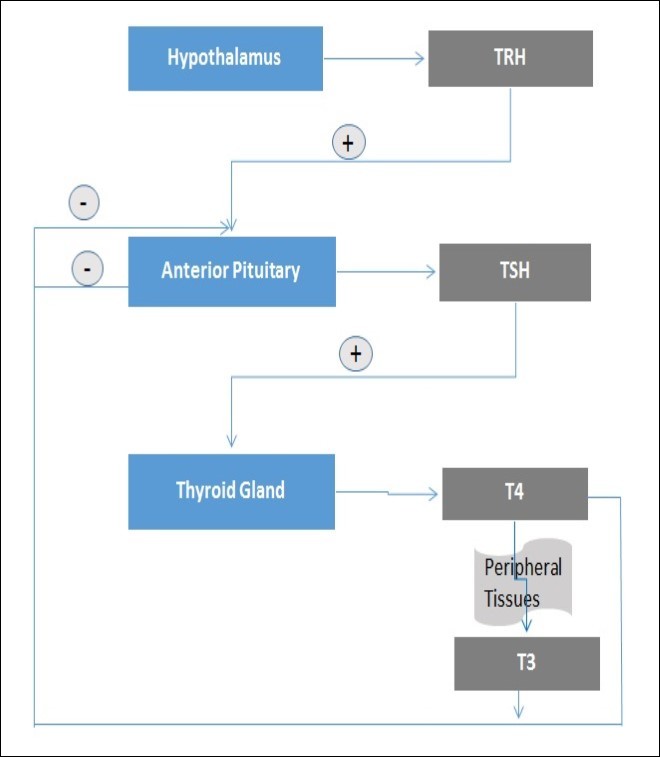
Download figure
Major factors that determine the body's energy need include resting basal metabolic activity, physical processes, BMI, nutrition frequency, and gender 32. This basal metabolic rate, an important parameter of energy requirement, is significantly associated to thyroid hormones. Moreover, there are evidences revealing thyroid hormone-dependent energy requirement is associated with adaptive thermogenesis 30, 33. The adaptive thermogenesis in question here defines the balance between heat loss and gain. The heat requirement in this equilibrium is associated with oxidative phosphorylation related to locally produced thyroid hormones in brown adipose tissue, which is more pronounced in some animals (such as the bear). And what’s more, thyroid hormones are partially effective on the energy requirement of the skeletal muscle 34. The fact that thyroid hormones promote increased energy expenditure during rest may be considered a function preventing the storage of energy as fat.The detection that adaptive thermogenesis due to thyroid hormones affects the energy expenditure during rest 35, 36, strengthens this idea. This relationship between obesity and thyroid functions contributes to the progress of researches in this direction.
Deiodinase Activity in Obesity and Chronic Diseases
Under normal conditions, 3,5,3’,5’-tetraio do-L-tyronine (T4) is synthesized more than 3,5,3’-triiodo-L-tironine (T3) in the thyroid gland. The majority of T3 in the blood is formed by removal of iodine from the outer ring of T4 with the 5’-deiodinase enzymes commonly found in peripheral tissues other than thyroid. While the less common 5-deiodinase enzymes by catalyzes the separation of iodine from the inner ring of T4, providing the formation of inactive reverse T3 (rT3). These deiodinase enzymes are classified into three types (D1, D2 and D3) according to their activity and synthesis in different organs. D1 is mainly expressed in the liver and shows its predominant activity on the outer ring of T4. The potent inhibition of D1 by anti-thyroid drug propylthiouracil (PTU) is an important feature that differentiates D1 from other deiodinases. It has been reported to be active in D2, central nervous system, pituitary, brown adipose tissue and skeletal muscle that can show its effect only on the outer ring. The expression of D2 is aimed at preserving tissue T3 levels against plasma levels of T4 and T3, which vary depending on the changes in thyroid function. D3, which is mainly expressed from the brain, has only the inner ring deionization activity and mediates the degradation of thyroid hormone. Therefore, brain tissue is considered to be the main site for the plasma T3 to be transformed by inactivation (cleaning) and plasma rT3 production 37, 38, 39, 40, 41. TSH and 5’-deiodinase enzyme activity may be suppressed when basal metabolic rate decreases in cases, which metabolism slows down (prolonged fasting, calorie restriction and some chronic diseases such as renal and liver failure), thus, T3 formation from T4 in peripheral tissues may be inhibited. In some studies, it has been reported that a relatively large amount of rT3 is formed in fetus, chronic diseases, carbohydrate starvation and diabetes mellitus 42, 43, 44, 45. Based on these findings; obesity, which is commonly seen in chronic diseases, will probably be negatively affected by the transformation of T4 to T3 since D2 is suppressed (Figure 2). Therefore, more rT3 formation will occur. However, more conclusive evidence is needed.
Figure 2. The 5’ and 5-deiodinase enzyme activity in obesity, prolonged fasting and chronic diseases.
Download figure
Important in Energy Metabolism of Thyroid Hormones
It is known that thyroid hormones play an important role in energy metabolism and alter the functions of other endocrine glands. The effect of thyroid hormones on insulin secretion function of the pancreas is not known yet, but its association with diabetes is detected. Studies on thyroid hormones focused on fat degradation, glucose oxidation and oxidative phosphorylation acceleration and other metabolic effects. In particular, thyroid hormones-mediated increased energy metabolism and insulin function have been highly emphasized. Recent studies suggest that thyroid hormones play an important role in the islet pathology of diabetes and have non-genomic effects on kinases or calmodulin, Ca+ 2-ATPase, adenylate cyclase and glucose transporters. However, it is assumed that T3 shows its most important effect through the regulation of thyroid receptors-mediated via target gene transcription found in the nucleus 46, 47, 48. Studies in adult rat have shown that T3 plays an important role in coordinated regulation of lipogenesis and lipolysis, basal oxygen consumption and storage of fats 49, 50. For all these reasons, the use of thyroid hormones in the treatment of obesity and diabetes continues to be a current topic.
Although there are studies indicating that there are elevated levels of leptin in parallel with fat ratios in hypothyroid rats and that thyroid hormones affect serum leptin concentrations, there is no conclusive evidence explaining the mechanism of relationship between leptin and hyper / hypothyroidism 51, 52, 53, 54, 55.
Many studies have reported that the increased leptin levels in rats with hypothyroidism are observed, and thyroid hormones affect serum leptin concentrations. However, there is no conclusive evidence defining the mechanism of the relationship between leptin and hyper / hypothyroidism.
Thyroid Hormone Levels in Obesity and its Contribution to ATP Consumption
In our study conducted in obese patients, we observed that there is a strong correlation between the severity of nonalcoholic fatty liver disease (NAFLD) and insulin resistance (IR) and blood TT3, fT3 and adiponectin levels. In this study, IR and NAFLD severity increased, especially fT3 levels decreased and then increased. We considered the possible cause of this last increase as the body's protection mechanism against severe pathology 56.In a recent study 34 reporting moderate TSH elevation (T3 is normal or slightly above normal) in obese individuals supports our finding.
Clinical studies have indicated that there are many causes of moderate TSH elevation in obese people. These reasons; a) subclinical hypothyroidism due to iodine deficiency or autoimmune thyroiditis, b) dysfunction of the hypothalamic-pituitary axis, c) thyroid hormone resistance and d) an adaptation process to increased energy consumption 57, 58, 59, 60, 61. In a large-scale cohort study, moderate levels of TSH elevation were found in many obese patients without any thyroid disease, but it was reported that some of the obese children and adults participating in the study had autoimmune thyroiditis 62, 63.
T4 release and blood T4 levels are rapidly suppressed during fasting. This suppression contributes to the adaptive reduction of the metabolic rate helping preservation of energy and reinforcing survival. The mechanism underlying this response has been associated with the dramatic reduction of circulating leptin during fasting. Prevention of the decrease in thyroid hormone levels associated with systemic administration of leptin during fasting 64, 65, this confirms the relation.
Considering the effects of thyroid hormones on metabolism, the relationship between obesity and thyroid hormone will be better understood. That is, the effect of T3 on Na+/K+-ATPase is well known 66 and T3 increases the ATP consumption by accelerating the activity of Na+/K+-ATPase in almost all tissues. Accelerating oxidative phosphorylation leads to the use of energy as heat instead of converted to ATP, as well as the synthesis of ATP. Therefore, O2 consumption and metabolic rate will increase 67. Thus, it will be ensured that the excess energy is consumed instead of storing it as fat.
Thyroid hormones increase the sensitivity of β-adrenergic receptors to catecholamines and then stimulate respiration, heart rate and systole (Figure 3). To achieve this, the rate of glucose metabolism through glycogenolysis and gluconeogenesis will increase. In animal studies, prolonged administration of T3 has been reported to stimulate GLUT4 expression and glucose transport in the skeletal muscle 68. It is thought that thyroid hormones can regulate glucose metabolism, and this is achieved through the glucose transporter (GLUT-4) via the T3-sensitive control point. In a clinical study, it was reported that the TCA cycle (without altering the rate of ATP synthesis) was accelerated by short-term administration of T3 to healthy individuals, accelerating the transport of glucose in skeletal muscles by T3-effective separation 69.
Figure 3. Specific functions of thyroid hormones on target organs.
Download figure
Under normal conditions, thyroid hormones stimulate protein synthesis (Figure 3). In the presence of excess thyroid hormone, protein synthesis is slowed down and negative nitrogen balance is observed 70. While thyroid hormones stimulate lipid synthesis in a similar way to protein, it is also reporting that it increases the activity of hormone-sensitive lipase at the same time. It is decreasing fat deposits by increasing fatty acid oxidation to form ATP for use in thermogenesis 71. Overproduction of thyroid hormones has been associated with hyperthermia, tachycardia, sweating and hypertension. However, heart rate deceleration, drowsiness, weight gain, constipation, high cholesterol level, slowing the basal metabolism rate, cold intolerance and psycho-somatic disorders are attributed to decreased thyroid hormone production 72.
Conclusion
Similar to leptin behavior, thereby it is understood that thyroid hormones increase in advanced stages of obesity and increase energy consumption at rest. Accelerating oxidative phosphorylation, with ATP synthesis, is thought to be the use of energy that is not transformed into ATP as heat, so it can be consumed instead of storing excess energy as fat. In addition, T3 hormone in the skeletal muscles without changing the rate of ATP synthesis of glucose transport and the TCA cycle of finding that the thyroid hormones may be an effective tool to stand against obesity. However, reporting that 5’-iodinease activity is suppressed in chronic diseases causes anxiety about in the efficacy of thyroid hormones in obesity. The increasing tendency of thyroid hormones in the advanced stages of obesity is probably a rescuer mechanism to increase the effectiveness of suppressed thyroid hormones.
As a result, in the advanced stages of obesity, increased in thyroid hormones supports to enhance energy consumption of the body by increasing the energy need during rest, thus offering an alternative rescuer way to the body against obesity risk. This situation is a rescuer adaptive contribution similar to the impact that leptin, which correlates with the amount of adipose tissue in obesity, inhibits feeding by affecting the satiety center.
Disclosure
There is no authors’ potential conflict of interest in this paper. The authors received no financial support for this study.
References
- 2.Dina C. (2008) New insights into the genetics of body weight. , Curr Opin Clin Nutr Metab Care 11, 378-384.
- 3.Reaven G, Abbasi F, McLaughlin T. (2004) Obesity, insulin resistance and cardiovascular disease. Recent Prog Horm Res. 59, 207-223.
- 4.Shamseddeen H, Getty J Z, Hamdallah I N, Ali M R. (2001) Epidemiology and economic impact of obesity and type 2 diabetes. , Surg Clin North Am 91, 1163-1172.
- 5.Jacobi D, Stanya K J, Lee C H. (2012) Adipose tissue signaling by nuclear receptors in metabolic complications of obesity. , Adipocyte 1(1), 4-12.
- 6.Zhang Y, Proenca R, Maffei M, Barone M, Leopold L. (1994) Positional cloning of the mouse obese gene and its human homologue. , Nature 372, 425-432.
- 7.Bruun J M, Lihn A S, Verdich C, Pedersen S B, Toubro S et al. (2003) Regulation of adiponectin by adipose tissue-derived cytokines: in vivo and in vitro investigations in humans. , Am J Physiol Endocrinol Metab 285, 257-533.
- 8.Romanatto T, Roman E A, Arruda A P, Denis R G, Solon C et al. (2009) Deletion of the tumor necrosis factor-α receptor 1 (TNFR1) protects against diet-induced obesity by means of increased thermogenesis. , J Biol Chem 284, 36213-36222.
- 9.Hotamisligil G S, Shargill N S, Spiegelman B M. (1993) Adipose expression of tumor necrosis factor-alpha: direct role in obesity linked-insulin resistance. , Science 259, 87-91.
- 10.Patel L, Buckels A C, Kinghorn I J, Murdock P R, Holbrook J D et al. (2003) Resistin is expressed in human macrophages and directly regulated by PPAR gamma activators. , Biochem Biophys Res Commun 300(2), 472-476.
- 11.Delaigle A M, Jonas J C, Bauche I B, Cornu O, Brichard S M. (2004) Induction of adiponectin in skeletal muscle by inflammatory cytokines: in vivo and in vitro studies. , Endocrinology 145(2), 5589-5597.
- 12.Berner H S, Lyngstadaas S P, Spahr A, Monjo M, Thommesen L. (2004) Adiponectin and its receptors are expressed in bone-forming cells. , Bone 35(4), 842-849.
- 13.Hedbacker K, Birsoy K, Wysocki R W, Asilmaz E, Ahima R S. (2010) Antidiabetic effects of IGFBP2, a leptin-regulated gene. , Cell Metab 11(1), 11-22.
- 14.Levi J, Gray S L, Speck M, Huynh F K, Babich S L. (2011) Acute disruption of leptin signaling in vivo leads to increased insulin levels and insülin resistance. , Endocrinology 152(9), 3385-3395.
- 15.Mehta S, Farmer J A. (2007) Obesity and inflamation: A new look at an old problem. Curr Atheroscler Rep. 9(2), 134-138.
- 16.Fain J N, Madan A K, Hiler M L, Cheema P, Bahouth S W. (2004) Comparison of the release of adipokines by adipose tissue, adipose tissue matrix, and adipocytes from visceral and subcutaneous adipose tissues of obese humans. , Endocrinology 145(5), 2273-52282.
- 17.Campfield L A, Smith F J, Guisez Y, Devos R, Burn P. (1995) Recombinant mouse OB protein: evidence for a peripheral signal linking adiposity and central neural networks. , Science 269(5223), 546-549.
- 18.Deem J D, Muta K, Ogimoto K, Nelson J T, Velasco K R et al. (2018) Leptin regulation of core body temperature involves mechanisms independent of the thyroid axis. , Am J Physiol Endocrinol Metab 315(4), 552-564.
- 19.Coppari R, Bjorbaek C. (2012) Leptin revisited: its mechanisms of action and potential for treating diabetes. Nat Rev Drug Discov. 11(9), 692-708.
- 20.MEF Vazquez-Vela, Torres N, Tovar A R. (2008) White adipose tissue as endocrine organ and its role in obesity. , Arch Med Res 39(8), 715-728.
- 21.Loffreda S, Yang S Q, Lin H Z, Karp C L, Brengman M L. (1998) Leptin regulates proinflammatory immune responses. , FASEB J 12(1), 57-65.
- 22.Konstantidines S, Schafer K, Koschnick S, Loskutoff D J. (2001) Leptin-dependent platelet aggregation and arterial thrombosis suggests a mechanism of atherothrombotic disease in obesity. , J Clin Invest 108(10), 1533-1540.
- 23.Turer A T, Khera A, Ayers C R, Turer C B, Grundy S M. (2011) Adipose tissue mass and location affect circulating adiponectin levels. , Diabetologia 54(10), 2515-2524.
- 24.Appel S J, Harrell J S, Davenport M L. (2005) Central obesity, the metabolic syndrome and plasminogen activator inhibitor-1 in young adults. , J Am Acad Nurse Pract 17(12), 535-541.
- 25.Siklova-Vitkova M, Klimcakova E, Polak J, Kovacova Z, Tencerova M. (2012) Adipose tissue secretion and expression of adipocyte-produced and stromavascular fraction-produced adipokine vary during multiple phases of weight-reducing dietary intervention in obese women. , J Clin Endocrinol Metab 97(7), 1176-1181.
- 26.Ma L J, Mao S L, Taylor K L, Kanjanabuch T, Guan Y. (2004) Prevention of obesity and insulin resistance in mice lacking plasminogen activator inhibitor 1. , Diabetes 53(2), 336-346.
- 27.Festa A, D'Agostino R Jr, Tracy R P, Haffner S M. (2002) Elevated levels of acute-phase proteins and plasminogen activator inhibitor-1 predict the development of type 2 diabetes: the insulin resistance atherosclerosis study. , Diabetes 51(4), 1131-1137.
- 28.Joseph-Bravo P, Jaimes-Hoy L, Uribe R M, Charli J L. (2015) 60 years of neuroendocrinology: TRH, the first hypophysiotropic releasing hormone isolated: control of the pituitary-thyroid axis. , J Endocrinol 226(2), 85-100.
- 29.Mullur R, Liu Y Y, Brent G A. (2014) Thyroid hormone regulation of metabolism. , Physiol Rev 94(2), 355-382.
- 30.Cioffi F, Gentile A, Silvestri E, Goglia F, Lombardi A. (2018) Effect of Iodothyronines on Thermogenesis: Focus on Brown Adipose Tissue. Front Endocrinol (Lausanne). 9-254.
- 31.Krotkiewski M. (2002) Thyroid hormones in the pathogenesis and treatment of obesity. , Eur J Pharmacol 440(2), 85-98.
- 32.Plucker A, Thomas D M, Broskey N, Martin C K, Schoeller D. (2018) Adult energy requirements predicted from doubly labeled water. , Int J Obes (Lond) 42(8), 1515-1523.
- 33.Iwen K A, Oelkrug R, Brabant G. (2018) Effects of thyroid hormones on thermogenesis and energy partitioning. , J Mol Endocrinol 60(3), 157-170.
- 35.Silva J E. (2003) The thermogenic effect of thyroid hormone and its clinical implications. , Ann Intern Med 139(3), 205-213.
- 36.Silva J E. (2006) Thermogenic mechanisms and their hormonal regulation. , Physiol Rev 86(2), 435-464.
- 37.Eugene J Barrett. (2012) The thyroid gland. In: Boron WF, Boulapep EL, eds , Philadelphia, PA, USA 1044-1056.
- 38.R Arrojo E Drigo, Bianco A C. (2011) Type 2 deiodinase at the crossroads of thyroid hormone action. , Int J Biochem Cell Biol 43(10), 1432-41.
- 39.Tu H M, Kim S W, Salvatore D, Bartha T, Legradi G. (1997) Regional distribution of type 2 thyroxine deiodinase messenger ribonucleic acid in rat hypothalamus and pituitary and its regulation by thyroid hormone. , Endocrinology 138(8), 3359-3368.
- 40.Guadano-Ferraz A, Obregon M J, St Germain DL, Bernal J. (1997) The type 2 iodothyronine deiodinase is expressed primarily in glial cells in the neonatal rat brain. , Proc Natl Acad Sci U S A 94(19), 10391-10396.
- 41.Peeters R P, Visser T J. (2000) Metabolism of Thyroid Hormone. In: De Groot LJ, Chrousos G, Dungan K, Feingold KR, Grossman A, Hershman JM, Koch C, Korbonits M, McLachlan R, New M, Purnell J, Rebar R, Singer F, Vinik A, eds.SourceEndotext[Internet].South Dartmouth(MA):MDText.com,Inc.;2000-.2017Jan1
- 42.Martinez B, Ortiz R M. (2017) Thyroid Hormone Regulation and Insulin Resistance: Insights From Animals Naturally Adapted to Fasting. Physiology (Bethesda). 32(2), 141-151.
- 43.Araujo R L, de Andra, de Figueiredo ÁSP, da Silva ML, Marassi M P. (2008) Low replacement doses of thyroxine during food restriction restores type 1 deiodinase activity in rats and promotes body protein loss.J.Endocrinol.198(1):. 119-125.
- 44.O’Mara B A, Dittrich W, Lauterio T J, DLS Germain. (1993) Pretranslational regulation of type I 5’-deiodinase by thyroid hormones and in fasted and diabetic rats. , Endocrinology 133(4), 1715-1723.
- 45.Takeuchi Y, Suzuki D, Oda S I, Refetoff S, Seki K. (2006) Unique regulation of thyroid hormone metabolism during fasting in the house musk shrew (Suncus murinus, Insectivora: Soricidae). , Gen Comp Endocrinol 146(3), 236-241.
- 46.Damiano F, Rochira A, Gnoni A, Siculella L. (2017) Action of Thyroid Hormones, T3 and T2, on Hepatic Fatty Acids: Differences in Metabolic Effects and Molecular Mechanisms. , Int J Mol Sci 18(4), 744.
- 47.Yen P M. (2001) Physiological and molecular basis of thyroid hormone action. , Physiol Rev 81(3), 1097-1142.
- 48.Chen C, Xie Z, Shen Y, Xia S F. (2018) The Roles of Thyroid and Thyroid Hormone in Pancreas: Physiology and Pathology. , Int J Endocrinol 2018-2861034.
- 49.Oppenheimer J H, Schwartz H L, Lane J T, Thompson M P. (1991) Functional relationship of thyroid hormone-induced lipogenesis, lipolysis, and thermogenesis in the rat. , J Clin Invest 87(1), 125-132.
- 50.Oppenheimer J H, Schwartz H L, Mariash C N, Kinlaw W B, NCW Wong. (1987) Advances in our understanding of thyroid hormone action at the cellular level. Endocr Rev. 8(3), 288-308.
- 51.Escobar-Morreale H F, F Escobar Del Rey, G Morreale De Escobar. (1997) Thyroid hormones influence serum leptin concentrations in the rat. , Endocrinology 138(10), 4485-4488.
- 52.Leonhardt U, Gerdes E, Ritzel U, Schafer G, Becker W. (1999) Immunoreactive leptin and leptin mRNA expression are increased in rat hypo- but not hyperthyroidism. , J Endocrinol 163(1), 115-121.
- 53.Pinkney J H, Goodrick S J, Katz J, Johnson A B, Lightman S. (1998) Leptin and the pituitary-thyroid axis: a comparative study in lean, obese, hypothyroid and hyperthyroid subjects. , Clin Endocrinol 49(5), 583-588.
- 54.Zimmermann-Belsing T, Dreyer M, Holst J J, Feldt-Rasmussen U. (1998) The relationship between the serum leptin concentrations of thyrotoxic patients during treatment and their total fat mass is different from that of normal subjects. , Clin Endocrinol 49(5), 589-595.
- 55.Syed M A, Thompson M P, Pachucki J, Burmeister L A. (1999) The effect of thyroid hormone on size of fat depots accounts for most of the changes in leptin mRNA and serum levels in the rat. , Thyroid 9(5), 503-512.
- 56.Ozcelik F, Yuksel C, Arslan E, Genc S, Omer B. (2013) Relationship between visceral adipose tissue and adiponectin, inflammatory markers and thyroid hormones in obese males with hepatosteatosis and insulin resistance. Arch Med Res. 44(4), 273-80.
- 57.Reinehr T, G de Sousa, Andler W. (2006) Hyperthyrotropinemia in obese children is reversible after weight loss and is not related to lipids. , J Clin Endocrinol Metab 91(8), 3088-3091.
- 58.Reinehr T, Isa A, G de Sousa, Dieffenbach R, Andler W. (2008) Thyroid hormones and their relation to weight status. Horm Res. 70(1), 51-57.
- 59.Mantzoros C S, Ozata M, Negrao A B, Suchard M A, Ziotopoulou M. (2001) Synchronicity of frequently sampled thyrotropin (TSH) and leptin concentrations in healthy adults and leptin-deficient subjects: evidence for possible partial TSH regulation by leptin in humans.J Clin Endocrinol Metab. 86(7), 3284-3291.
- 60.Kok P, Roelfsema F, Langendonk J G, Frolich M, Burggraaf J. (2005) High circulating thyrotropin levels in obese women are reduced after body weight loss induced by caloric restriction. , J. Clin. Endocrinol.Metab 90(8), 4659-4663.
- 61.Kozlowska L, Rosolowska-Huszcz D. (2004) Leptin, thyrotropin, and thyroid hormones in obese/overweight women before and after two levels of energy deficit. , Endocrine 24(2), 147-153.
- 62.Reinehr T, Hinney A, G de Sousa, Austrup F, Hebebrand J. (2007) Definable somatic disorders in overweight children and adolescents. , J. Pediatr 150(6), 618-622.
- 63.M1 Rotondi, Leporati P, La Manna A, Pirali B, Mondello T et al. (2009) Raised serum TSH levels in patients with morbid obesity: is it enough to diagnose subclinical hypothyroidism?. , Eur J Endocrinol 160(3), 403-408.
- 64.Blake N G, Eckland D J, Foster O J, Lightman S L. (1991) Inhibition of hypothalamic thyrotropin-releasing hormone messenger ribonucleic acid during food deprivation. , Endocrinology 129(5), 2714-2718.
- 65.Ahima R S, Prabakaran D, Mantxoros C, Qu D, Lowell B. (1996) Role of leptin in the neuroendocrince response to fasting. , Nature 382(6588), 250-252.
- 66.Everts M E. (1996) Effects of thyroid hormones on contractility and cation transport in skeletal muscle. , Acta Physiol Scand 156(3), 325-333.
- 67.McAninch E A, Bianco A C. (2014) Thyroid hormone signaling in energy homeostasis and energy metabolism. , Ann N Y Acad Sci 1311, 77-87.
- 68.Zorzano A, Palacín M, Gumà A. (2005) Mechanisms regulating GLUT4 glucose transporter expression and glucose transport in skeletal muscle. , Acta Physiol Scand 183(1), 43-58.
- 69.Lebon V, Dufour S, Petersen K F, Ren J, Jucker B M. (2001) Effect of triiodothyronine on mitochondrial energy coupling in human skeletal muscle. , J Clin Invest 108(5), 733-737.
- 70.Mondal S, Raja K, Schweizer U, Mugesh G. (2016) Chemistry and Biology in the Biosynthesis and Action of Thyroid Hormones. Angew Chem Int Ed Engl. 55(27), 7606-7630.
Cited by (1)
This article has been cited by 1 scholarly work according to:
Citing Articles:
Al- Anbar Medical Journal (2022) OpenAlex Semantic Scholar