Abstract
One of the key steps in determining how to prevent the viral disease is to identify the virus. The virus lives in different ways and in different environments. It lives in the air, in the sea, on plants, animals and objects and humans. Some people put humans on the path of developing zoonotic diseases that are specific to animals but also involve people with unhealthy behaviors. In the food chain, each animal is hunted by other animals and feeds on other animals or plants and other objects. Bacteriophages are viruses that kill bacteria. And there are creatures that kill viruses and this is the biological struggle with viruses. When the virus enters the body, it performs a series of activities that lead to a series of symptoms in the patient. These symptoms include the behavior of viruses. These are among the ecological and behavioral characteristics of viruses that need to be fully understood in order to limit viruses and deal with epidemics and pandemics. In this study, we try to reach a conclusion by reviewing the articles that have information about the behavioral (signs and symptoms) and ecological characteristics of viruses and use these findings in order to combat viruses.
Author Contributions
Academic Editor: Raul Isea, Fundación Instituto de Estudios Avanzados -IDEA, Venezuela.
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2020 Baratali Rezapour, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Competing interests
The authors have declared that no competing interests exist.
Citation:
Introduction
In late December 2019, a series of unexplained cases of pneumonia were reported in Wuhan, China. The government and health researchers in China have taken swift steps to control the spread of the epidemic and have launched an etiological study. On January 30, 2020, the WHO declared the Coronavirus-2019 an emergency public health epidemic concern (PHEIC). On February 11, 2020, WHO officially named Coronavirus-2019 (COVID-19) a disease caused by 2019 novel coronavirus. On the same day, the International Group on the Classification of Viruses by the Coronavirus Study Group (CSG) named as the Severe Acute Respiratory Syndrome (SARS-CoV-2). Several classes of patients with unknown pneumonia in Wuhan, Hubei Province, China, have been reported to Chinese health authorities since December 8, 2019, and most of these cases were epidemiologically related to the local fish and animal market. The pathogen responsible for these pneumonia classes was identified as a 2019-nCoV. At the beginning of the 2019-nCoV outbreak in China, it remained much unknown, except for the fact that it was transferred to the market with direct exposure. However, person-to-person transmission has been confirmed since 2019-nCoV and asymptomatic individuals have been identified as a potential source of infection. The number of identified cases has been steadily increasing, and as of February 3, a total of 14,557 cases have been reported worldwide. Since the first laboratory-confirmed case was identified on January 20, 2020 in Korea, the number of reported cases has increased to 15 since 3 February 20201. Following the SARS crisis, awave of structural proteomics pervaded the coronavirus research community. The unusual pro-proteomic side of SARS at the time was the focus on proteins with specific roles in pathogenesis, and rival international teams tried to solve structures and determine functions throughout the viral proteome 2. the WHO has stated that to date, no specific drug has been recommended for the prevention and treatment of COVID-19 in China, but historically, when the spread of the disease began, Chinese medicine was used, including oral prescription herbal formulas, the use of Chinese medicines (CM), and indoor medicinal plants. In 2003, for example, the CM method was used to prevent and treat acute respiratory distress syndrome (SARS), the most serious infectious disease in China before COVID-19. In 2009, during the H1N1 flu epidemic around the world, the National Directorate of Traditional Medicine of China issued a CM prevention program that included four Chinese herbal medicinal formulas (CHM) for adults with different CM compounds and another for children. The current prevalence of COVID-19 has led many provinces in China to issue prevention and control programs for CM, including prevention programs, mainly the oral CHM formula3. Virusologists have traditionally focused on viruses that cause disease in humans, domestic animals, and crops, but recent advances in the metabolic sequence, especially the high-power sequence of peripheral specimens, have viruses. Remarkable has been revealed everywhere on the planet. There are at least 1031 virus particles at any given time in the world in most environments, including marine and freshwater habitats and metastatic digestive systems, in which the number of detectable virus particles is 10 to 100 times the number of cells. In addition to their significant abundance, viruses are remarkably diverse in nature and the organization of genetic material, gene sequences and encrypted proteins, and the mechanisms of reproduction and interaction with their cellular hosts, whether antagonistic, or reciprocal. Although dsDNA viruses that kill bacteria (bacteriophages) are best studied, recent research shows that about 50% of marine viruses have the ssDNA or RNA genome. In fact, a large number of complete or almost complete genome sequences have been collected from metagenomic data for viruses with small, medium and even large genomes. Identifying completely new groups of viruses from such analyzes, emphasizing the power of metagenomic approaches to virus detection, some of which can be key functions in setting up ecosystems, while others can coexist with their hosts without causing disease recognizable or even reciprocal. Recently developed tool VirSorter has identified about 12498 new viral genome sequences in 15,000 bacterial and archaeal genomes, which have increased the number of known prokaryotic viruses by almost 10 fold and identified viruses that infect 13 prokaryotephyla. Only by accepting sequences produced by metagenomic methods do they really represent existing viruses, including placing them in classification schemes, we can hope to better understand the ecology, history, and impact of the global virus4. Type I IFNs play a crucial role in the formation of antiviral immune responses signaling through type IFN receptors leads to the activation of a specific set of genes, including kinase R protein and Mx proteins. They have direct anti-viral effects. Gene-stimulated IFN-type I products, such as gamma IFN, activate the downstream elements of the innate immune system that further clear viral pathogens, although almost all hematopoietic cells And non-optopathies are able to produce disinfection after viruses, but pDCs are the main source of alpha IFN in both humans and mice (an important feature of pDCs). TLR7 and TLR9 receptors are essential for viral pathogen sensation and induce innate immune response, respectively. In cytomegalovirus infection (MCMV), pDCs respond rapidly and the first wave IFNs produce alpha. Previous studies have clearly shown the role of pDCs in the rapid production of type IFNs in antiviral immune responses. In this study, we developed a unique function. Here are a few basic facts about a stomp pad and how it is used. In addition, we have identified identified pDCs as the source of human IFN non-response to SARS-CoV, which plays an important biological role in IFNs of the pDC-derived type for highly pathogenic coronavirus infections in humans5. Disruption of the communication network and CNS regenerative properties make this organ vulnerable to microbial agents as well as physical injuries. Therefore, there is little endogenous antigen in the CNS or the potential to activate T cells. Limited expression of adhesion molecules by endothelial cells of the blood-brain barrier and strong bonds between these cells also limit or prevent the entry of large molecules such as antibodies and T cells into the CNS. However, several active cells or CD4 + and CD8 + memory randomly patrol the CNS in the absence of "danger" signals, and exit or die if the antigen is not detected. In addition, several monitoring mechanisms limit the immune response to prevent damage to uninfected host cells or to induce autoimmunity (viruses that remain in the human CNS include DNA viruses that are exemplary). It is the herpes simplex virus and JC polymovirus RNA viruses such as measles virus; HIV and HTLV-16.
Coronaviruses infect many species of animals, including humans. Molecular mechanisms, replication, and pathogenesis of several coronaviruses have been actively studied since the 1970s. Some animal viruses, such as Transmissible Gastroenteritis Virus (TGEV), Bovine Coronavirus (BCoV), and Infectious Bronchitis Viruses (IBV), are of veterinary importance. MHV has been studied as a model for human disease. The family of viruses is relatively obscure, probably because there is no serious human disease that can be definitively attributed to coronavirus. The human coronavirus only causes colds. However, in the spring of 2003, when it became clear that a new human coronavirus was responsible for SARS, coronaviruses became much more widely known. With the spread of SARS, coronaviruses may be considered "emerging pathogens." Since the SARS epidemic, two new human coronaviruses have been described. In 1975, the Coronaviridae family was founded by the International Committee on Virus Classification. Recently, at the 10th Nidovirus International Symposium in Colorado Springs, Colo In June 2005, the Coronaviridae family was proposed to be divided into two subfamilies, coronaviruses and toroviruses, the latter of which cause intestinal diseases in cows and possibly humans. The Coronaviridae families along with the Arteviridae and Roniviridae families make up Nidovirales. The Arteviridae family includes pig and horse pathogens, and the Roniviridae family is composed of vertebrate viruses. Coronaviruses are divided into three genera, commonly known as the group, and based on the serological interaction; the analysis confirmed the sequence of the newer genome of this group. Coronaviruses are found in the first group of terrestrial animal pathogens, such as TGEV pigs, Porcine Epidemic Diarrhea Virus (PEDV), and FIPV, as well as Human Coronaviruses (HCoV 229E) and HKU1 transplants that cause respiratory infections. The second group includes veterinary-related pathogens such as BCoV, swine angioglobulin encephalomyelitis virus and horse coronavirus, as well as human OC43 and NL63 coronaviruses, which, like HCoV-229E, cause respiratory infections. The second group also includes viruses that infect mice. The sialodacryoadenit coronavirus also belongs to the second group. We have listed SARS-CoV in the second group. The third group so far includes only poultry coronaviruses, such as IBV, turkey coronavirus, and pheasant. Recently, using RT-PCR, coronavirus sequencing in gray goose and wild pigeons was detected. In February 2003, the World Health Organization received reports from China on the prevalence of a new respiratory disease in Guangdong Province was cultivated. Sequential information indicates that this was a previously known coronavirus. The virus has been shown to be pathogenic for SARS through infections in non-human mammals. The SARS epidemic was officially controlled until July 2003. The epidemic was controlled only by severe isolation of patients at the end of the epidemic, the CDC and WHO reported more than 8,000 deaths, more than 800 worldwide. SARS cases were reported in 29 countries, mostly in Asia, although North America was also affected, the most important being Toronto, Canada. However, only 8 confirmed serological evidence for SARS-CoV infection. Coronaviruses are an attractive group of viruses that offer pathogenic animal models, unusual molecular transcription and recombination mechanisms, and emerging pathogens. The emergence of SARS and the identification of a coronavirus as the causative agent of the disease were surprising to the coronavirus community, as it was the first significant and effective association of coronavirus with severe human disease. Although SARS-CoV will re-emerge among the human population, it has encouraged awareness that coronaviruses may be the cause of human respiratory disease and possibly other types of disease. The identification of NL63 and HKU1 shows examples of newly described human sizes. The data collected over the years make it possible to research animal crown crowns, identify SARS-CoV very quickly, and sequence genomes. Knowing that multiple viral genes are involved in pathogenesis, and especially in the type of immune response, tells us that small changes in sequence can have a greater impact on the pathogenic phenotype. Observations that coronavirus tropical species may be easily selected during reproduction in tissue culture or animals, and different species selected by changing the region created in different regions in tissue culture medium, are all useful in understanding the emergence of SARS in the human population are identification and characterization of proteases and replication, as well as the identification of several enzymatic activities coded in ORFs 1a and 1b. The experience of developing coronavirus vaccines will also contribute to the development of SARS vaccines. Future paths for SARS-CoV research include further understanding of reproduction mechanisms7.
Coronaviruses are a family of viruses that can cause a wide range of diseases, from colds to SARS. In November 2002, a severe and deadly respiratory illness caused by a new coronavirus in China was identified, killing 8098 patients and 744 deaths due to SARS in one year. The deadly respiratory syndrome was called SARS. Since June 2003, the World Health Organization has not reported any cases of human infection with the virus. In 2012, a Qatari patient contracted a severe respiratory illness, and microbiological studies showed that the cause of the disease was a new virus from the cocoronavirus family and the SARS-Like virus. A study of the respiratory patterns of dead Jordanian patients showed that the first cases of infection and death of patients due to a new deadly virus called nCoV occurred in Jordan (6 months earlier than Qatar) significant point in the epidemiology of coronavirus. The new high mortality rate compared to SARS, which had about 10 percent mortality, was the highest mortality in SARS syndrome in the age group over 65 years (about 50 percent). It is known for humans. Previous experience with SARS syndrome has shown that the lowest mortality rate was in the under-24 age group (1%)8. Most infectious diseases in poultry are caused by RNA viruses. Their specific characteristics, such as high mutations, short production times, and large populations, help these viruses to evolve rapidly. Coronaviridae are now divided into two subfamilies, Coronavirinae and Torovirinae. According to the latest ICTV update, coronaviruses are classified into four genera: Alphacoronavirus, Betacoronavirus, Gammacoronavirus, and Deltacoronavirus. This finding may indicate that wild birds may carry viruses similar to IBV for free, forcing us to monitor gamma-ray coronavirus monitoring in quail farms9. FCoV has recently been designated as a member of subgroup 1a in the Coronaviridae family. Other members of the subgroup include TGEV, CCV, RDCoV and CFBCoV10. In a few short years, the dramatic increase in the number and variety of new virus genomes discovered through metagenomic methods in various environments, from marine to terrestrial, from tropical forests to wastewater, from hot springs to Antarctic lakes, is understandable. It has changed us from the global virus in general and the RNA virus in particular. Far from the trivial suffixes of previous knowledge derived primarily from the comparison of plant and animal viruses, new metagenomic studies change the existing picture of virus evolution. In addition, these new discoveries appear to indicate a close and intricate connection between the evolution of the virus and the evolution of the host environment that was previously unclear. These changes are made to existing ideas about the evolution of the virus primarily in three areas. First, the proliferation of many virus groups provides stronger genes for protected phylogenetic genes (primarily, RdRps, in the case of RNA viruses). Eliminating suspicious clusters of RNA viruses such as foliage and the like has replaced the stronger branch of tombus-noda, which demonstrates the similarity of genome architecture among the smallest and simplest RNA viruses. However, fundamental changes have been made by large groups of previously identified viruses, such as Picorna and Alpha-like viruses, although they generally lose some environmental members, but the test appears to be very varied he does. Second: the interplay of gene modules among the various genomes of the virus, previously known as an important evolutionary process in the pre-metabogenic period, increases dramatically with metaganomic discoveries. Combined Tombus-noda viruses and alpha-aseptic viruses such as TMV capsid protein coding can be cited as some notable examples. This is a predominant trend in the evolution of viruses with different types of genomes that have been shown, especially with the discovery of the huge variety of Kimmeric ssDNA viruses. Recent metagenomic studies suggest that RNA viruses are not backward in this regard. Third, and most notably, the inclusion of extensive metabogen data in phylogenetic analyzes has led to the identification of multiple and highly supported groups that combine RNA viruses that infect a variety of hosts, including different groups of proteins, animals, and plants. Of course, these observations come with serious precautions, including possible contamination. Phylogenetic artifacts, such as the absorption of long branches, and Dolja are probably incorrect samples of viruses from different host species. These warnings, however, show that the continuous combination of viruses from different hosts in phylogenetic tree branches throughout a wide range of RNA viruses reflects the reality of virus evolution. The main aspect of this fact is the wide range of hosts of many virus groups, which is why HVT appears to be a determining factor in the evolution of the RNA virus. The major role of HVT in virus evolution varies between virus groups and hosts. In particular, the RNA virus is very diverse from the plant RNA virus dense by picorna-like, alpha-like, tombus-like, partiti-like, reo-like, bunya-like, rhabdo-like, and some other RNA virus clades. It is crowded. Such a massive RNA virus can be caused by the vertical descent of the ancestral family of Zygnematophycea algae or HVT from the invertebrate virus much more extensively or through a combination of these pathways. It seems that the small algae taxon of the ancestors hosts all the viral components in the flowering plant. In addition, the algae virus is known to be dominated by Phycodnaviridae, a large DNA virus that has been completely eliminated from the plant virus. Therefore, although some components of the plant virus can lurk in the Zygnematophycea, this hypothesis of plant RNA virus growth through HVT from the marine environment through freshwater, soil and air routes is preferred. Such HVTs are likely to be mediated by nematodes, aquatic animals, and terrestrial inhabitants, which are the most abundant animals on Earth, arthropods, and, perhaps to a lesser extent, mollusks and fungi. A similar argument can be made for vertebrates, albeit with less certainty: At present there is no information about viruses Deuterostomes other than vertebrates, while Protostomes viruses, in particular, arthropods and nematodes, are very diverse. Along with the well-known vector characteristics of many arthropods and nematodes, this makes the HVT of these animals the most likely pathway for the development of the vertebral virus, according to current data. However, proper sampling of echinoderm viruses, vertebrate groups, as well as animal proteins, such as Choanoflagellates, is necessary to support or rule out this line of reasoning. The role of HVT in the evolution of some RNA viruses has already been suggested, for example in the case of the plant RNA virus (-) RNA, whose ancestors appear to have been derived from animals, but the pervasive feature of this phenomenon has been revealed by metagenomics. It represents a change in the sea of our understanding of the evolution of the RNA virus. In particular, the diverse RNA virus from invertebrates, which temporarily encompassed both vertebrates and angiosperm plants, was a reservoir that created vertebrate and plant viruses. The role of invertebrates in the evolution of the virus is consistent with the lifestyle of many arthropods and nematodes. Aside from vector transmission, HVT is clearly facilitated in aquatic environments, compared to terrestrial environments, where viruses appear to be confined to a wide variety of hosts, such as various proteins and animals. Accurate tracking of possible HVT events is problematic due to possible changes in the evolutionary rate after the host change. However, in many cases, it appears that HVT is clearly involved in the evolution of the host and the global environment, such as the Cambrian Animal Explosion or the landing of animals and vascular plants. The new metaviromic data appear to support three main pathways through which eukaryotic RNA viruses have evolved. In the first case, a continuous line of descent from RNA (+) bacteriophages was obtained by viruses such as Narna, the RdRp of which is derived from Leviviridae, and some of which are still produced in mitochondria. Leviviridae and some of them still reproduce in mitochondria. The second pathway, as exemplified by Picorna viruses, involves the genome assembling of the ancestral virus from genes derived from bacteria, their viruses, and selfish elements. Although the ultimate evolutionary power of RdRp, like Picorna, has not been fully established, it appears to have evolved from RT within the bacteria of the second bacterial group. The third route involves the recent origin of major groups of viruses due to the lack of evidence for RNA (-) viruses, flu-like viruses, and alpha-like viruses in eukaryotes in invertebrate vertebrae have emerged, late evolution at the root of Metazoa. As an in-depth sampling of invertebrate RNA viruses, all three pathways appear to be extensive during a wide variety of aquatic invertebrates using the invertebrate RNA panvirome. Many invertebrates are excellent HVT agents, because of their close biological connections to organisms such as vertebrates and plants; they often act as vectors that keep viruses alive between these organisms. Despite the importance of invertebrates, they could not remain alone in the seeds of existing aquatic and terrestrial viruses. The first manifestations of the fungal virus indicate that it is richer than previously expected. Because fungi are ubiquitous organisms whose population size can be compared to or greater than invertebrates, in-depth sampling of fungi for viruses could potentially account for a large proportion of fungi in Show RNA formation of eukaryotic pan virus. To develop such a great plan, at least two major advances are needed: first, large-scale phylogenetic analysis of highly divergent proteins, such as RdRps and RT, and second, "evolutionary virus," a regular study of all viruses the main species of the host. In addition, it is likely that a more metagenomic compound, especially with the most powerful sequence-like detection methods, will lead to the discovery of completely new groups of viruses in what is now the "dark matter" of RNA metagenomes. Such findings could significantly alter the evolution of the RNA virus11.
Viruses are the most abundant microbes in the sea, most likely the entire biosphere. In addition, they have emerged as important geochemical and environmental factors in marine ecosystems. The metagenomic virus appears either specifically by deep sequencing of enriched environmental samples for virus particles or by identifying specific virus sequences in databases from other metagenomic projects. Marine viruses are genetically the most diverse biological communities on earth. The main and most unexpected finding was that the vast majority of viral sequences were not significantly similar to any sequences in the current database, and these sequences, which have traceable homologues, indicate that primarily, different bacterial genes often play an important role in central metabolism instead of separate classes of genes commonly found in known bacteriophages or other viruses. These significant findings suggest two possibilities that are not mutually exclusive. First, known viruses may not represent real viruses, for the reason that marine viruses are the main reservoir of new genes in the ocean. Second, specimens that appear to be representative of viruses may not be primarily of viral origin and reflect sample contamination with non-viral DNA, indicating a serious defect in current metagenomic protocols. Here, we use several computational methods to analyze marine dsDNA viruses and show that, despite the negligible contamination with bacterial genes, these sequences represent a set of statistical features. Genomically, prokaryotic and known viral are quite different. Therefore, there seems to be a realistic possibility that true marine viruses are composed mainly of virus-like particles that are different from the marked phages and may be similar to gene transfer agents. Sequence-like searches have shown that at least 50-60% and, typically, close to 90% of the resulting DNA are read proteins that are significantly similar to others encoded in known genes or viruses or cellular origins are not encrypt. This low percentage of sequences with recognizable similarities to known viral proteins may indicate that many, or perhaps most, metabolic sequences show new virus genes that have no adaptation in databases because of the actual diversity of viruses. If a substance separated by enrichment of viral particles is in fact composed of GTA-like components, it is fair to conclude that metagenomics is changing the ideas in the structure and dynamics of the prokaryotic world. Biospheres, viruses have repeatedly sampled the whole variety of bacterial genes. Recent advances in virus metabogens have shown that viruses and quasi-virus elements are the most abundant biological organisms on the planet, and that their genomic diversity is inferred from model systems. In addition, the analysis of metagenomic data suggests a distinct possibility that the predominant forms in viruses may be qualitatively different from those identified and may be similar to GTAs. The implications of these findings are that gene transfer pathways between the virus world and the "sustainable" genome of cell life forms could be much broader than previously thought. The metagenomics virus has had other unexpected findings, such as the discovery of a large number of sequences apparently caused by the eukaryotic DNA viruses NCLDVs in marine metagenomes, which are apparently dominated by prokaryotes and an almost unique display of viruses such as Picorna in RNA marine viruses. Despite these findings, the study of virus diversity in the environment, and in particular viral metagenomics, is clearly in its infancy, and advanced technologies are needed to provide sample and sequential data analysis. Some of the most likely ways of future research include the following: a complete description of marine viruses (in which proteomics will play an important role). In addition to studying new viruses from a variety of habitats, especially severe ones, a thorough analysis of the role of viruses in the environment and geochemistry and estimating the true size of the Korean virus, to adequately reconstruct the evolution of the entire virus world, must be metagenomic approach with census. Completed viruses infected with all major hosts of prokaryotic and eukaryotic hosts. There is no doubt that with the advent of new generations of proteomic sequences and technologies, the metabolic virus has a bright future and will significantly help in the emerging new understanding of the genetic diversity of life12.
The Origin of the New Coronavirus-2019
Epidemiological studies of primary cases of novel coronavirus-2019 pneumonia have shown that many cases have been exposed to the Hanan seafood market in Wuhan, China. It has been identified from the Hanan seafood market. Thirty-three of the 585 environmental samples available in the Hanan seafood market were positive for COVID-19. Some include live animals, such as hedgehogs, zebras, snakes, turtles, birds, and possibly pangolins, but do not include bats. Therefore, bats are not likely to come into direct contact with humans, and direct transmission of the virus from bats to humans seems unlikely because, in the past, although SARS-CoV and MERS-CoV originated from bats as the main and natural reservoir, they were transmitted to humans through hosts of a certain type of cat (civet) and more. The novel Coronavirus-2019 is a chimeric virus between the bat coronavirus and the coronavirus of unknown origin. Compared to other animals, they found that snakes are most likely the reservoir of the novel Coronavirus-2019. The COVID-19 is very closely related to the coronavirus isolated from a particular species of Chinese bat (chrysanthemum-headed) and this virus is highly associated and high homology with the bat-bison coronavirus. Thus, the novel 2019 Coronavirus may have originated in bats and then been transmitted to humans through an intermediate host in the Hannan market. Currently, the novel 2019 Coronavirus has been isolated from anteaters (pangolins), and it has been found that the strains isolated from humans infected with the novel 2019 Coronavirus are 99% similar. It has been suggested that the route of evolution and evolution of the novel Coronavirus-2019 was from bat to anteater (intermediate hosts) and then to humans13. A wide variety of viruses have been similarly reported from insects and several eukaryotic and prokaryotic viruses have been identified in terrestrial environmental samples. Metagenomic studies have also discovered many surprising viruses in the human gastrointestinal tract that have not been diagnosed before despite decades of research. For example, the 97-kb dsDNA bacteriophage genome, called crAssphage, is six times more abundant in the general metagenomic data set than the sewage sample or sewage effluent than other known bacteriophages. The virus accounts for up to 90% of the readings read in metagenome derived from virus-like particles and makes up 1.7% of the total human fossil metagenomic sequence read in public databases14.
Serological and genetic evidence from various studies supports the genomic origin of SARS-CoV. This hypothesis was first shown by epidemiological reports that early patients with SARS in Guangdong Province were exposed to live wild animals in markets that traded restaurants. In order to identify animals carrying SARS-CoV, a wide range of domestic and wild mammals in Guangdong Province were examined. Interestingly, viruses similar to SARS were genetically linked to all human SARScoronavirus use using RT-PCR in the shepherds' noses and feces of Nyctereutesprocyonoides cats. Serological evidence of infection was observed in these species as well as in a deer Melogale moschata. Interestingly, animal traders working with live animals in these markets had a high prevalence for human and animal SARS virus, although they did not have a history of the disease. SARS-like animals isolated from animals were more than 99% similar to human SARS-CoV. However, compared to animal viruses, the SARS human virus isolated from the early stages of epidemics and showed deletions in ORF 8 from May 2003 that vary in length (from 29 to 82 nucleotides in the early stages) led to a 415 nucleotide deletion in the loss of the entire ORF 8 region at the late stage of the prevalence. It is not clear whether elimination in ORF 8 is beneficial to humans or whether ORF 8 can be spread in humans but not in animals. The SARS coronavirus animal precursor may not be deficient in human infection, and exposure to the leading animal virus may lead to an abortion infection or antigen stimulation. The serological response observed in animal ranchers. Therefore, live animal markets have probably been a place for animals to transmit the virus to humans. In this sense, molecular epidemiological studies have suggested the introduction of several animal coronaviruses in humans. For example, phylogenetic analysis of the few human cases that occurred in December 2003 in Gong Dong Province showed that this SARS-CoV is much closer to the CoV civet palm than the SARS-CoV isolated from humans in the primary epidemic. Whether SARS-CoV has a reservoir in one species of wild animal remains to be seen, In addition, domestic cats are prone to SARS-CoV infection. Transmission occurred when non-infected cats came in contact with an infected animal, and viral headlines gradually increased, the authors reported. Interestingly, the efforts of several groups to identify wildlife reservoirs led to the discovery of novel animal coronaviruses in bats and birds. Meat goat coronaviruses, first-class dogs, and cats are linked to antigens, and may even range from a common ancestor. To support this idea, coronavirus infection may occur in pigs, dogs, and cat animals. In addition, dogs and foxes may be used as reservoirs for TGEV. The discovery of ACE2 as a SARS-CoV receptor is a major breakthrough in our understanding of how SARS-CoV enters cells. The two groups compared the protein content. Protein S represents a mild prevalence from 2003 to 2004, and one of the palm civets mediates the more effective infection of ACE2-expressing cells than human-expressing cells. In contrast, protein S is effectively linked to the severe outbreaks of 2002 to 2003 and uses both receptors. This data is consistent with the lack of human-to-human transmission during the 2003-2004 outbreaks and the recent transfer of SARS-CoV to humans. This difference in the ability of human civet and SARS isolates to use human receptors is associated with amino acid exchange in RBD. There is disagreement about the SARS-CoV classification. Early in its discovery, SARS-CoV was proposed to create the fourth new group among coronavirus groups. Stavrinides and Guttman15 reported the possibility of recombination between parent viruses such as mammals and birds. The authors proposed a mammalian origin such as replication protein, bird origin such as matrix and nucleococcal protein, and the mosaic origin of birds from the S protein. However, the origin of SARS-CoV with recombinant mammalian and poultry viruses seems unlikely. While recombination is one of the hallmarks of coronavirus and there is evidence that it is probably one of the main forces in the evolution of coronavirus, there is no evidence of recombination among members of different coronavirus groups. Accordingly, using chimeric recombinant viruses, it has been shown that the 3’ UTR SARS-CoV may replace a function for this MHV, whereas 3’ UTRs from the first prototype groupI or the second coronavirus group cannot16. Therefore, the evidence to date suggests that SARS-CoV belongs to the second group of coronaviruses7.
Alphacoronaviruses and Betacoronaviruses are isolated from several mammal species, including humans, dogs, cats, and cows. However, all known Gammacoronaviruses only infect bird species with some exceptions examples include infectious bronchitis virus (IBV), turkey coronavirus (TCoV), and pheasant corona. Infection, meanwhile, has been reported in several other species, including Greylag geese, Mallard ducks, pigeons, and quail. These findings, along with the separation of Gammacoronaviruses from several other species of birds, have led experts to suspect the role of some species, such as quail, as CoV reservoirs and carriers of CoV on IBV epidemiology9. Phylogenetic analyzes showed that the sequence of M and 7b genes in viruses derived from healthy cats and those derived from diseased cats showed a coexistence of both biotypes in cats10. The discovery of previously suspected RNA viruses in the sea has dramatically expanded our knowledge of the ecology of the global virus and led to more in-depth research on the origin and evolution of Picorna viruses. Recently, it has been found that the phylogeny of viruses such as Picorna and their host are radically unpleasant. The most plausible explanation for this lack of intervention is that, at least in the early stages of their evolution, viruses such as Picorna do not empathize with hosts, but before irradiating superconducting eukaryotic groups with 5 original host’s evolved viral diversity. In addition, the main genes of viruses such as Pycnura were transmitted from bacteria, recycling bacteria and phages to ancestors. The richness of the marine community of viruses such as Picorna is in stark contrast to the lack of other marine RNA viruses. To date, only one RNA-positive virus from quasi-fluvial families of the virus and none of the families such as the alphavirus family has been identified in environmental samples. An important explanation for the dominance of a virus such as Picorna could be a limited scale of RNA metabolism studies, which may be in a biased representation of the diversity of the virus in the environment. On the other hand, the representation of marine RNA viruses in current metabolic samples may be sufficient, meaning that single-celled eukaryotes can actually be infected primarily by Picorna-like viruses. Such dependence on RNA viruses such as Picorna with single-celled eukaryotic host hosts appears to be the ancestral group from which RNA-positive eukaryotic multicellular eukaryotic viruses evolve. Extensive sampling of marine RNA viruses has the potential to test this hypothesis, and perhaps, marine specimens were used from all major evolutionary breeds of positive and animal RNA viruses. The metagenomic study of the evolution of positive RNA viruses updates the eukaryotic strand, but the origin of the two-strand and negative RNA viruses remains unknown. To date, only a few double-stranded, virus-like viruses and no RNA viruses with negative strains have been identified in single-celled eukaryotes, while a wide variety of viruses in both classes infect animals, including marine invertebrates and vertebrates it is obvious. Both bipolar and negative RNA viruses are common in terrestrial plants; however, neither class is as diverse as RNA viruses. This global environmental model is compatible with at least two possibilities. First, double-stranded and negative RNA viruses may have emerged following the emergence of multicellular eukaryotes from positive or retrograde RNA viruses, possibly in several relatively recent independent cases. Second, RNA viruses are less likely to be bipolar and negative than their ancient eukaryotes, but for unknown reasons, they are now endangered or limited to sheds that have not yet been sampled by metagenomics. However, a more in-depth study of marine RNA viruses is likely to provide evidence for this prominent problem of virus evolution12.
Viral Features
Viral features that can be traced from tracking data, including genome organization, reproduction strategy, presence of homologous genes and, potentially, host domain or vector type, may be additional biological characteristics. Such information is better than genomic sequencing, which includes the potential for complete coding of the virus, and should be at least classified based on sequences. Viruses that have a genomic sequence and are not closely related to viruses in existing species pose a particular problem because there is no phenotypic standard that can be classified. For example, bacteriophage classification is typically based on the sequence and structure of virions, but these characteristics may be used to classify RNA viruses in animals and plants whose deeper relationships are often in RNA gene polymerase sequences and other repetitions. Protected is obvious, not appropriate4. West Nile virus and encephalitis virus primarily target nerve cells. Other human viruses, herpes viruses, are targeted and hidden in nerve cells. The HIV and JC viruses primarily target other CNS cells and are prone to latent or persistent CNS infections6. Coronaviruses are an "RNA virus". These viruses are more likely to be genetically mutated, and the more likely they are to remain in the human population, the more likely they are to develop an unpleasant, malignant mutation8. Mutations and recombination in the CoVs genome have led to viruses with different tissue tissues, increased vigor, and increased stamina in chickens. The viruses were phylogenetically distinct from the infectious bronchitis virus, and all Gammacoronavirus isolated in quail were different from IBV vaccines. If the vaccine-like strains were similar, viruses were 85.74-86.64 and 85.16-86.47 percent, respectively, similar to the 91.4 and H120 vaccine species. From 2016 to 2018, 47 herds were examined in which 4 positive herds (8.5%) were located in Tehran province9. According to the phylogenetic analysis of the N gene, the viruses were clustered in a separate group other than the known Gammacoronaviruses. The least similarity (24/97%) between ACov / Quail / UT-BPG2 / 2017 and ACov / Quail / UT-BPG3 / 2017 and the highest similarity (100%) between ACov / Quail / UT-BPG3 / 2018 and ACov / Quail / UT-BPG4 / 2018 Separations were most similar to IR-Ur1_09 (34/87 34) and IS_1618_07 (88.59%). All positive samples were also positive for the UTR gene9. Thus, SV is highly enriched for specific FAG genes, although they make up only a small proportion of the total genes in the sample (<0.5%) and a relatively small proportion of FOG specific POGs (<10%)17. In contrast, sequences in SV correspond to a significantly lower proportion of protected cellular orthologs compared to STf, and are much less likely to have the best results with known prokaryotic proteins. Enrichment of viral proteins despite the presence of normal bacterial genes in SV is indescribable, for example, by comparing the content of the coding sequences of ribosomal components between SV and STf. They do not encrypt, and reliance on host ribosomes to express the genome is included in most modern definitions of viruses. In addition, comparative genomic data suggest that ribosomal protein genes, despite several proven exceptions, are susceptible to any type of horizontal gene transfer. Therefore, the virus metagenomas analyzed from cell gene markers are depleted compared to non-viral metagenomas, but SV and OV contamination with non-cellular gene genes are negligible. Unlike DNA viruses, RNA viruses are microscopic and difficult to detect with an epilepsy microscope, so the relative prevalence of RNA viruses in the environment is still poorly known. The sequence of RNA-derived viral genomes is also not insignificant because RNA is fragile and needs to be reversed in DNA. Apart from all these problems, RNA metabolism has already had a major impact on our current understanding of the evolution of the virus and global ecology12.
Structure of Novel Coronavirus 2019 (COVID-19)
Coronaviruses are enveloped, non-segmented viruses that have a single-stranded RNA, a Positive-sense of animal origin, and belong to the Coronaviridae family and the Nidovirales category (Figure 1). The size of the virus genome is between 26 and 32 kb, which is one of the largest RNA viruses. These viruses have two different types of surface proteins and are named after this apparent feature. The coronavirus family is genotypically and serologically divided into four sexes: alpha, beta, gamma and delta. About 30 species of coronaviruses have been identified in humans, mammals and birds. Human coronaviruses are caused by alpha and beta. Coronaviruses are among the most common viruses, with 30 to 60 percent of China's population having antibodies against them18. COVID-19 is the third known zoonotic animal disease known after SARS and MERS respiratory syndrome, both of which belong to the beta-carotene category19.
Figure 1.Source CDC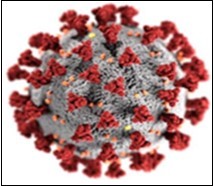
The first two parts of replicase, nsp1 and nsp2, are somewhat obscure, but they appear to directly support virus replication by intervening with host defense. The nsp3-6 subunits include all the viral factors that are essential for the formation of repetitive viral organelles20, as well as the two proteins that are responsible for processing all viral replication proteins21. Small subsets of nsp7–11 include viral initiation activities and basic support for reproduction 22, 23, 24. The last part of the replicase of nsp12-16 contains the residual RNA correction enzyme for replication and correction. The replication organization has a kind of chronological term. Nsp1–2 helps colonize the host, followed by Nsp3–6, which forms the basis for organizing and protecting replicas. This follows nsp7–11 manufacturing activities that also interact with outpatient payments and RNA synthesizers25. Finally, in the right context, RNA-synthesizing enzymes from C-trminal replication are able to function. Replicase proteins are produced from large proteins and are therefore produced at the same time. For this reason, the order in which different proteins are active during the viral replication cycle is not poorly understood. The replication organization also follows almost the same sequence26, 27.
Epidemiological Characteristics of COVID-19
From January 10 to 24, 2020, the number of people infected with the Novel Coronavirus-2019 infection in China increased 31.4 times. On February 23, 2020, the number of people living with COVID -19 in China in 1879 was 10 January 2020. The death-to-death ratio for women was 3.25 to 1, the median age of death was 75 years, the median time from the first symptoms to death was 14 days, and the median time from early symptoms to death was 70.5 days shorter than those under 70 years of age (20 days). COVID -19 may progress more rapidly in adults than in young adults. The rate of transmission of the disease from the infected person was 2.2. Although 55% of the first patients infected with the Novel Coronavirus-2019 were related to the Hanan seafood market, the number of unrelated cases has increased exponentially (logarithmically) since late December 2019. 73% of patients with coronavirus infection was male and 32% of patients had underlying diseases such as diabetes (8 patients), hypertension (6 patients) and cardiovascular disease (6 patients). The median age was 49 years. Of the 41 patients, 27 were associated with the Hanan seafood market. The mortality rate of patients with modern coronavirus-2019 was 15%. The rate of transmission of infected patients to individuals was 3%. The mortality rate of patients with novel coronavirus-2019 was 14%19. According to Wuhan, China estimates the number of reproductions 0.3 to 2.68. The average incubation period was 6.1 days and the average interval was 7.7 days. The average time to double was between 6.4 days and 7.7 days the mortality rate of hospitalized patients at 15-15%. We cannot identify any study that reported infectious and latent courses. The estimated number of reproductions of 0.3 was obtained from a small number of infected patients with incomplete information in the early stages of the outbreak, so the number of reproductions 2019-nCoV is likely to be similar to SARS 2002/2003 during the pre-intervention period (domain, 2 to 3) and influenza virus. Gear A / H1N1 was released in 2009 in the United States. The incubation period is likely to be similar to that of coronavirus SARS, but with a greater confidence interval (mean 4.8 days; CI 95%, 4.2 to 5.5). In addition, it is longer than the A / H1N1 pandemic influenza virus in 2009 (average incubation period, 1.4 days; CI 95%, 1.0 to 1.8). Therefore, the evidence examined above shows that current control measures for 2019-nCoV, including quarantine and observation time of 14 days for suspected cases, can be considered appropriate. Serial production time and distance 2019nCoV is longer than the A/H1N1 pandemic influenza virus in 2009 (average production time, 2.7 days; CI 95%, 2.0 to 3.5 and average serial distance: domain, 2.6 to 3.2). However, the mean interval between the 2019-nCoV series is similar to SARS (mean, 8.4 days; standard deviation, 3.8). The overall mortality rate in 2019-nCoV was estimated by international experts at 3% to 14%, and is more likely to cause infection in older age groups1. The Novel coronavirus pathogenes is mechanism of 2019 and the use of receptors while both FIPV and FECV may cause the virus only FIPV is repeated in macrophages and causes disease10.
The Life of the Virus
Coronaviruses bind to specific cellular receptors via the spike protein. This causes a change in spike and then mediates the fusion between the viral and cell membranes, that leading to the release of nucleo-capsid into the cell. At high frequency (25 to 30%) 25, 27, 28, 29, 30 Frameshift function occurs. The mechanism of processing pp1a and pp1ab into the mature was replicated proteins by encoding one or two papain-like proteases and a picornavirus 3C-like protease (by ORF 1a)31. An additional enzymatic activity, cyclic phosphodiesterase, is encoded downstream in ORF 2a. These enzymatic activities play a role in the role of RNA metabolism or interference with host cell processes. Infection with coronaviruses, like other RNA viruses, genome proliferation and mRNA transcription must occur. Genome proliferation involves the synthesis of all negative RNA strands that exist at low concentrations. D and is used as a template for complete genomic RNA. Subgenomic 3coterminal RNAs interact with each other, and full-length genomic RNAs act as mRNAs. Each mRNA has a common leader sequence at its 5 'end. In addition, negative-strand RNAs are related to the length of each mRNA as well as the full genomic length of the low-level. The mechanism of adaptation to the group of positive and negative RNAs that are made includes a unique case. It is individual and consists of mechanical structures that are not fully understood. However, it is believed that subgenomic mRNA synthesis with a transcription regulatory sequence, which is present in the RNA gene, is predicted at mRNA transcription initiation sites. The current model, in which discontinuous transcription occurs during the synthesis of negative RNAs associated with manganese, with antifreeze sequences at the end of the negative RNA 3s, which are then used as templates for the synthesis of mRNAs. Viral proteins from separate mRNAs are generally translated from 5 ORFs. In some cases, there may be two ORFs that have been translated from one mRNA. After translation, M and E membrane proteins are located in the Golgi intracellular membrane near the site. Therefore, reluctance to M, other viral or cellular factors may be needed to determine the location of the sprout. M and E proteins, expressed in the absence of other viral proteins and viral RNA, are sufficient to produce antiviral particles. Protein S is distributed over intracellular membranes as well as plasma membranes. Protein S interacts with the transmembrane protein region during assembly. For some viruses, cell-to-cell fusion occurs as a spikemediate, thus promoting syncytium. The protein complex of the nucleocapsid with the RNA genome forms spiral structures. Protein N interacts with protein M and germination of vesicles occurs. The virus then spreads to the surface of the cell, where it leaves the cell. Interestingly, TGEV and MHV exit the epithelial cells from the opposite side. When the two viruses are used to experimentally infect the same cells, the epithelial cells of the pork representing the recipient, TGEV is preferably released in the apical membrane, while MHV is preferably placed at the basal level that the vesicles containing the two coronaviruses are targeted. This indicates that the two viruses in Golgi are transmitted to different transport vesicles that have information that directs them to different levels. Therefore, differences in diffusion location may contribute to differences in virus propagation between TGEV and MHV. TGEV causes a localized intestinal infection, while MHV spreads to different organs7.
Natural Enemies of the Virus
Rapid Production of Type IFN in pDCs Following Infection MHV
In study “Control of coronavirus infection through plasmacytoid dendritic-cell–derived type I interferon” at the first set of this experiment, the IFN’s type response of pDCs and cDCs after exposure to MHV was determined. For this purpose, CD11clow B220 + PDCA-1+ pDCs and CD11c + B220 + cDCs were classified as splenic cell suspensions and contaminated with MHV. High IFN-a production in pDCs is well associated with viral infection control. Bone marrow-derived pDCs and cDCs, which were differentiated with Flt-3L or GM-CSF (growth factors), respectively, responded in a similar pattern: rapid and high production of IFN-a in pDCs (but not in cDCs) and a Well containment of viral replication by pDCs. A period of time RT-PCR analysis approved that type I IFN response in cDCs was significantly slower than pDCs5. In addition, pDCs that do not have type IFN receptors are more susceptible to MHV infection than wild-type pDCs. Therefore, it seems that pDCs are well able to respond effectively to MHV with a strong type IFN, and this initial reaction has an important protective effect against this cytopathic virus. The expression of TLRs that recognize viral products such as oligonucleotides CpG or ssRNA indicates that pDCs represent a highly specialized cell that provides the primary response to a specific set of infectious agents. Gives another feature of pDCs is quite high presentation of the IFN-7 adjustment factor-7 (IRF-7), which independently stimulates IFN-a expression directly from an intermediate IFN-b feedback loop. In MHV infection, this effective direct induction of IFN-a appears to be not only necessary to regulate of the magnitude of the IFN response type I, but also to limit the proliferation of this cytopathic virus in pDCs. In addition, type I pDC- derived IFN provide bystander effect protection because the primary reproduction of MHV in lymph nodes such as the spleen is reduced in the presence of pDCs. It is noteworthy that in MHV infection this function of pDCs cannot be replaced by other cells, for example, in MCMV infection. Slowly replicating viruses, such as MCMV, may not be able to fully demonstrate the importance of pDCs in controlling of cytopathic viruses that require a rapid response IFN typeI. In secondary lymph nodes, macrophages are the main target cell for MHV, and recent investigations show that cDCs can also be easily infected with MHV A59 or MHV JHM. A new study by Samit et al.32 shows that pDCs not only help minimize respiratory syncytial virus infection but also help create T-cell antiviral responses in the lungs. In addition, pDCs that do not have typeI IFN receptors are more susceptible to MHV infection than wild-type pDCs. Therefore, it seems that pDCs are well able to respond effectively to MHV with a strong typeI IFN, and this premature response has a strong protective effect against this cytopathic virus.
Type IFN Signaling is Necessary to Control MHV Infection
Through the IFN typeI receptor (IFNAR) signaling is essential for the control of several viral infections. To assess the importance of typeI IFN signaling during MHV infection, IFNAR- deficient mice (IFNAR _ / _) and 129Sv wild-type (wt) mice were infected with 5 pfu MHV. MHV infection in IFNAR _ / _ mice was lethal in only 48 hours, while wt mice survived without showing clinical disease associated with signs of MHV infection. In addition, IFNAR _ / _ but not wt mice were rapidly increasing the value of hepatic enzymes in the serum and an acute liver hemorrhage with extensive hepatic necrosis.
Careful periodic analysis of viral spread in both IFNAR _ / _ and wt mice showed that an IFN-type I functional system to limit the proliferation of primary virus to the spleen and prevent its spread in non-hematopoietic organs such as lung and central nervous system. It is essential that severe hepatotropic MHV proliferation in the liver in the presence of an IFN-typeI functional system is reduced by 3 to 4 times in terms of efficacy by reducing viral titles. Therefore, it is likely that this deadly disease rapidly develops in IFNAR _ / _ mice following MHV infection as a result of inadequate primary control of the cytopathic virus in the spleen and high levels of proliferation in various organs, ultimately leading to acute multiorgan failure.
Initial Control of MHV Infection Through Type I IFN Derived From Pdc
Primary MHV control requires an effective type IFN response that may be generated by pDC. PDCs use the TLR path instead of the RNA helicase RIG-I to identify RNA viruses and produce type IFN. Therefore, to study how MHV was detected by pDCs, bone marrow-derived pDCs from TLR3-deficient, TLR3 and TLR7 two eliminators (TLR3_ / _ / TLR7_ / _) TLR7-deficient, TLR7_ / _ and MyD88-deficient MyD88_ / _ mice were infected with low-dose of MHV (moi=1) and the prolifration of IFN-a was shown after 24 hours. Significant amounts of alpha IFN were observed in supernatant TDR3 - / - and wt control pDC culture. Therefore, to examine how MHV is detected by pDCs is triggered exclusively with the TLR7/ MyD88 pathway.
To assess the importance of alpha-derived IFN-derived IFN during MHV infection in the body, pDCs were eliminated using PDCA-1 antibody. As explained by Krug et al. For MCMV, a decrease in pDC was associated with a sharp decrease in serum IFN levels following MHV infection. Treatment with PDCA-1 resulted in 80% discharge of splenic pDCs for approximately 48 hours. However, it can have profound effects on viral headlines. Transient pDC reduction following low-dose MHV infection does not lead to mortality. However, the initial viral titers in the spleen were more than 1000-fold higher in pDC compared with the isotope-controlled mice, and the virus was found in other organs such as the lungs or brain. In order to eliminate global changes by supplementing the immune system, we evaluated the effects of reducing natural killer (NK) cells through the anti-Asian GM1. Decreased NK cells have neither altered the primary viral proliferation nor the IFN level in the serum. Finally, ALT levels in weak PDCA-1 mice have increased compared to control animals, indicating significant liver damage. These data clearly show that pDCs are important for the early control of MHV infection, and the absence of pDCs not only leads to the proliferation of uncontrolled viruses and spread to various organs, but also to the severity of viral disease.
Rapid Induction of Type IFNs in pDCs Following SARS-CoV Infection
In order to link the above to a human and potentially lethal human coronavirus infection, the ability of pDCs to produce alpha IFN after exposure to SARS-CoV was investigated. Primary pDCs and cDCs were isolated from the peripheral blood of healthy donors and infected with SARSCoV. As described for monocyte-derived cDCs, the main CDCs of healthy donors were also unable to produce significant amounts of alpha IFN and transcribed beta IFN genes and stimulating IFN genes such as ISG56 and MxA. On the IFN-type signaling pathway, it did not adjust regularly. In contrast, and as expected from MHV experiments, pDCs were able to produce alpha IFN early in SARS-CoV infection. In addition, mRNA expression for IFN beta, MxA, or ISG56 was found in infected pDCs. Based on this evidence and the unsuccessful attempts of previous studies to determine the type of IFN-producing cell in response to SARS-CoV, we conclude that pDCs are likely to be the main source of type IFNs in SARS-CoV infection5.
The first chemokines induced in the CNS seeking MHV infection were CXCL10 and CCL333. CXCL10 is expressed early in the first day by infected and disinfected glial cells. NK cells are absorbed by signal via CXCR3. Despite the rapid but transient absorption of NK cells in the CNS, there is little direct evidence for antiviral activity. However, their potential for IFN-γ secretion may facilitate the delivery of antigen by class I and class II MHC molecule regulation. CCL3 may enhance adaptive immune response by stimulating T cell activation and recruitment. Macrophages are the largest component of intrinsic CNS penetration. Their accumulation is increased by CCL5, which is induced by a slightly delayed kinetics compared to CXCL10 and CCL3. CNS infection with other neurological viruses, for example, lymphocytic choriomeningitis virus, TMEV virus, and measles, creates profiles expressing the chemokine gene-like MHV, indicating that CNS-resident cells are similarly infected and possibly from. They respond by expressing type I interferons. Quickly induced by MHV, cytokines are mainly present in astrocytes and microglia, including IL-1α, IL-1β, IL-6, and IL-12. Patterns similar to intrinsic cytokines, albeit with modified relative levels, are also characteristic of other CNS viral infections, including TMEV, vesicular stomatitis virus, HIV virus, and West Nile virus. This suggests that the secretion of these cytokines is a general antiviral or antiviral specific response and is consistent with their role in the subsequent activation of adaptive immunity. TNF-α, IL-12, and IL-1β mRNA levels increase, even in the absence of inflammation, indicating a CNS-responsive cell response to MHV infection. Induction of IL-6 plyotropic cytokines may enhance the passage of inflammatory cells throughout the BBB, similar to its role in the CNS autoimmune model, enhancing experimental allergic encephalomyelitis. T cells are ultimately unable to achieve sterile immunity or repress the virus, most likely due to a lack of regulation or inhibition of destructive constructive functions within the body. However, T cell cell dysfunction is complemented by a wave of virus-specific ASCs that are absorbed into the CNS following acute infection. In contrast to T cells, ASCs in the CNS are maintained at high frequencies during virus persistence. These data suggest that topical secretion of neutralizing antibodies in the CNS maintains the virus at low levels, thus preventing the virus from re-emerging by using an on-site protective system6.
The role of the immune response to MHV infection in virus clearance and pathogenesis in CNS has been well established. Both antibody and cellular immune responses are needed to protect against coronavirus infections. T cells CD8 + and CD4 + are primarily responsible for virus clearance during acute infection. Perfurin-mediated mechanisms are essential for virus clearance from astrocytes and microglia, while gamma interferon is involved in clearance of oligodendrocytes. It is not yet clear how the virus clears neurons. In the case of contamination with MHS-A59 from CNS, the transfer of specific epithelial CD8 + T cells prior to infection significantly reduces viral proliferation and the spread of viral antigens during acute infection, resulting in demyelination 4 weeks after infection decreases. This and other data show that the development of demyelination depends on the sufficient spread of the virus in the acute phase. While MHV is primarily cleared by the immune response through the cell, in the absence of B cells, antibodies are essential to prevent the virus from returning to the CNS after the T cell. Interestingly, this need for virus proliferation and clearance is not related to the liver. JHM neuromuscular infection is characterized by a strong and long-lasting response with alpha IFN, along with high levels of chemotherapy in macrophages such as CCL3, CCL4, CXCL2, asCXCL10, and CXCL57.
The New Coronavirus Pathogenesis Mechanism of 2019 and the Use of Receptors
Zhao et al34. found that ACE2 is a receptor for the new Coronavirus-2019. In the normal human lung, ACE2 is expressed in type 1 and 2 alveolar epithelial cells, with 83% of type 2 alveolar cells expressing ACE2. Men have higher levels of ACE2 in their alveolar cells than women. Asians have higher levels of ACE2 expression in their alveolar cells than white and African American populations. Connection of the new Coronavirus-2019 on ACE2 increases the expression of ACE2, which can lead to damage to alveolar cells. Injury to alveolar cells can in turn cause a number of systemic reactions and even death. They also confirmed that Asian men are more likely to develop coronavirus infection in 2019. Pre-attack of coronavirus on the host cell connects to the recipient. After binding to the receptor, the viral spike protein is broken down by catapsin-dependent protein proteolysis, TMPRRS2, or proprinase, and subsequently the viral coatings of cellular gardens are merged. The spike is a large, carnation-shaped trimmer that can be broken down by proteases into a subunit n S1 terminal that contains the second receptor junction (RBD) and a terminal S2c region. Compared to other coronavirus proteins, spike protein has the most variable sequence of amino acids, the strongest of all coronavirus genes to adapt to its hosts. Recent results show that the new Coronavirus-2019 from the same cell receptor ACE2 receptor, such as coronavirus sarce. In addition to the human ACE2, the new Coronavirus-2019 can use ACE2 honeycombs, a kind of cat (civet) and pig, but the ACE2 mouse cannot be used as an input receiver. One explanation is that the human ACE2 is similar Top with bat horses, Chinese horses, and cats, pigs 80.7%, 83.5% and 81.4%, respectively. The similarity of human ACE2 with anteater and cat is 84.8% and 85.2%, respectively. The similarity of human ACE2 with other ACE2 varies from 59.7% to 83.2%. In short, the new 2019 Coronavirus may have a wide range of hosts19. In several coronaviruses, nsp1 suppresses expression of the host gene. SARS-CoV nsp1 binds to the 40s subset and suppresses expression of the host gene. It has also been shown that SARS-CoV nsp1 mRNA destroys host but RNA species of coronavirus protect against degradation. A point mutation at the site of protein cutting between nsp1 and nsp2 in the complete TGEV genome releases nsp1 from the fledgling polyprotein and significantly reduces virus recovery. Komitani et al35, Have shown that the expression of nsp1 using plasmid greatly increases protein expression. This association is associated with reduced production at specific mRNAs, while rRNAs remain unpolluted. Overall, the transferred nsp1 mRNA gene, which was coated with polyadenylated, reduced host protein synthesis, and the combination of actinomycin D (to prevent new transcription) showed a much stronger inhibition of protein synthesis in the presence of nsp1. While translating new texts (cells are not treated with actinomycin D), translation of pre-existing versions was blocked by nsp1. Decreased mRNA levels and decreased preexisting mRNA translation were also likely to be seen as a result of degradation during SARS-CoV infection. SARS-CoV nsp1 has also been shown to be a potent stimulant of CCL5, CXCL10 and CCL3 expression in human lung epithelial cells activating NF- ĸB36 . The pathogenesis of SARS-CoV infection is characterized by a high immune reaction and a high increase in chemokine levels. In contrast, HCoV-229E, HCoV-OC43, and MHV do not significantly induce chemical deficiency, perhaps because they only cause mild upper respiratory tract disease. The nsp1 expression kinetics suggested that it may play a primary regulatory role in the viral life cycle. Nsp1 is the first adult protein to be processed from gene 1 polyprotein and is likely to be rapidly eliminated after translation of PL1pro nsp3. Mutated MHV, which is unable to release nsp1 from emerging polyprotein, is associated with delayed repetition, decreased peak titles, small plaques, and decreased RNA synthesis compared with wild-type control. These results emphasize the importance of nsp1 occurrence for optimal viral RNA synthesis and suggest that nsp1 may play an important role in MHV proliferative complexes. In MHV, nsp1 interacts with p10 and p15 (SARSnsp7 and nsp10 counterparts, respectively). Studies of immunolocalization and interaction in MHV have also shown that within the body, nsp1 may interact with other viral proteins - counterparts of SARS nsp2, 5, 8, 9, 12, 13 and sars9a37. However, at different times in the MHV life cycle, nsp1 has a different displacement than P65, the SARS counterpart nsp2. Studies of Y2H and Co-immunoprecipitation show that nsp1 interacts with E and sars3a. Nsp2 functions are unknown. In MHV, p65 plays an important role in the viral life cycle which appears to be distinct from its internal counterparts in coronaviruses. Based on immunoclonalization investigations in MHV, p65may done with SARS nsp1, 5, 7, 8, 9, 10, 12, 13 and sars9a counterparts38. Mutations with infectious clones SARS and MHV showed that nsp2 could be used for viral replication in cell culture. Always removing the nsp2 encoding sequence reduces virus growth and RNA synthesis. The exact nature of the role of nsp2 in viral growth and RNA synthesis is not yet clear. However, IBV nsp2 is a poor PKR antagonism, which may point to the complementary role of nsp1 in intracellular immunity. According to immunoassay studies in MHV, p65 may act in coordination with its counterparts SARS nsp1, 5, 7, 8, 9, 10, 12, 13 and sars9a. In 1993, Calver et al. showed that nsp3 may be involved in RNA synthesis of coronavirus. UB1 has a high structural similarity to Ras-interacting proteins. RAS family proteins (RFPs) act as clock molecules that rotate between inactive production and active GTP ranges. RFPs control cell growth, motility, and intracellular transport differentiation. Ras plays a key role in cellular prognosis from phase G0 to G1. Molecular interactions that lead to Ras inactivation prevent progression to G1. SARS-CoV and its other effects, such as MHV, are able to stimulate cells in G0 / G1phase during lytic infection cycles to their proliferative advantage. Sars3b plays a role in this process and nsp3 may also be involved in cell cycle arrest in G0phase. In addition, the UB1 is structurally similar to the ISG15. This protein combines with cellular targets as a major response to interferon alpha / beta, induction, and other markers of viral infection. High levels of this protein are essential for cellular antiviral response. It has been shown that ISG15 is able to prevent the spread of the virus by canceling nuclear processing of RNA precursors. However, some viruses have created a mechanism to prevent ISG15 expression. For example, the influenza B virus stops blocking by using the NS1 protein to overcome the immune response. It is possible that PL2prodomain nsp3 may bind to ISG15 and destroy the cell's antiviral response. NMR experiments showed a ligand limited to UB1, which was identified as a small RNA fragment by mass spectrometry. UB1 MHV has recently been shown to bind to nucleoproteins and effectively bind nsp3 to viral RNA during the proliferation process. This activity does not require a second AC to follow UB1 and is highly variable. SARS ADRP easily hydrolyzes 1 Phosphate group of Appr-1 -p is shown in laboratory conditions to be an active enzyme. Another group confirmed the finding: SARS ADRP and HCoV-229E human antibodies have been shown to show that Appr-1 -p to ADP-ribose to dephosphorylate to DEP-ribose in particular, that the enzyme lacks tracking activity in several Phosphate is another nucleoside. The role of ADRP in the coronavirus life cycle may be closely compared with the eukaryotic tRNA degradation pathway. Egloff et al. 39 suggested that ADRP could initial be a poly-ADP-ribose binding module. PARylation takes place in compromised cells for trigger apoptosis. PAR polymersses (PARPs) are responsible for labeling proteins. PARP activates DNA recognition and helps repair DNA. It activates automatically PARylates iteself, in the event of severe DNA damage, and destroys its nucleotide pool cell. If ADRP binds PAR, then PARylated proteins such as PARP attach to it. In fact, the second binding may be more beneficial because it can intensify the protein, apoptosis, prevent nucleotide depletion, and prolong virus replication and transcription in the infected cell. Due to the presence of ADRP in CoV nsp3s due to its important role in the viral life cycle is argued. Its function as ADRP in organic phosphate recycling seems to be a playable function and is not related to satisfaction with this domain. It appears that its role as ADRP may play an important role, as does its proposed role as a .PAR module. SUD-NM tends to have rich sequences of G and G squares, while SUD-MC showed the overall priority of purine nucleotides. While the SUD-N and SUD-M domains are structurally similar to the SARS-CoV ADRP domain, none of the domains have any significant dependence on ADP-ribose. This cluster is made up of three SARS-CoV nsp3 macroadenomas through gene proliferation, and SUD-M helps nsp3 function as an adjunct to the virus process. Comparing the activity of deo-coitinase between the wild and mutated Ubp6 species that do not have the Ubl domain, these associations are responsible for 300 times the increase in illegal speed and serve to activate the enzyme. PLpro-like domains act as a kind of "bait" or "deception" to neutralize cellular enzymes from other viral proteins, or they may interact with proteins between replication components. The physiological significance of its excitability in the viral replication cycle is not yet fully understood. However, structural E protein is easily absorbed into cells infected with ubiquitin, indicating that a reduction in the rate may be important in the assembly process. There is currently evidence that PL2pro is created by transcription activation pathways, by disabling TBK1, blocking NF-kappaB signaling, and preventing IRF3 transmission. NAB has been shown to contract as a hemodialysis at 37. C and has shown high strength for nucleic acid. While NAB was able to interact with single-stranded, double-stranded nucleic acids, it released single-stranded RNA nucleic acid protein cooling, indicating that NAB may act as an RNA-like binding ssRNA. So nsp3 is likely to perform a key role in a variety of hosts. UB1and SUD and RBD connect to RNA; also ADRP is a part of RNA processing machinery. If proteinases are not present, nsp3 is classified exclusively as RNA binding / modifying protein. These areas have been shown to alter nsp4 localization and cause membrane replication phenotype in transfected cells. If nsp3 participates directly in the membrane displayed in cells transferred with SARS-CoV nsp3 and nsp4, then the ZF domain is likely to participate in this deletion. Nsp4 is an essential component for the formation of vesicles of two viral membranes. It has also been shown that Nsp4 interacts with nsp2 on a two-hybrid yeast plate as well as with other nsp4 molecules in cells. Nsp4 has been shown to cause undetected DMV detection and lead to the elimination of nsp4 glycosylation. And become LC3-II. MHV Nsp6 shifts when it is expressed as nsp4, indicating that the two proteins interact with each other. It has also been shown that40 Nsp6 interacts with nsp2, nsp8, nsp9 and sars9b by measuring two yeast hybrids41.The bizarre observations provided convincing evidence that nsp8 is specifically associated with a molecular switch consisting of a cleft palate ring and pseudoknot RNA that is present in region 3 untranslatable MHV. Collaborates nsp9 may protect the fledgling ssRNA from nucleases during virus RNA synthesis due to its natural abundance in the cell. While the exact role of nsp9 in viral replication is not yet clear, Minkis and colleagues who examined the role of dimerinterface showed that SARS-CoV nsp9 is essential for effective viral growth. The first task was performed to perform nsp10 on the threshold of MHV species that contain temperature-sensitive lesions that synthesize viral RNA. It was further stated that this defect in nsp10 could not compensate for nsp4 or nsp5 in cells affected by virus-infected lesions, and this suggests that coronavirus 1a polyprotein is at least one unit from nsp4 onwards. It constitutes an important application that is negatively intermittent. Mutation studies have confirmed the importance of nsp10 for general RNA synthesis and for controlling the subgenomic ratio to genomic RNA. Deleting nsp10 or resetting the nsp7-10 genes encoding, fully translated of the Mpro site between nsp9 and nsp10 decrease virus growth42. Using financial and unexpected services, this budget has a temperature-sensitive lesion in nsp10 with the responsibility of Mpro services to have a transfer permit43. This way we can edit using nsp10 with the RNA tradition. The following Nsp10 can be reported from the mRNA fraudulent methylation complex to be edited is discussed below for the subtypes of viral methyl transferase25. RdRp is the central enzyme in the complex of several virral polypase compounds that replicates the viral RNA genome, including several other viral thinners. RdRp is the central enzyme in the multi-component viral replicase complex that replicates the viral RNA genome and includes several other viral thinners. Nsp13 is a helix that is able to ignore both RNA and DNA duplexes in the '5 to 3' direction with high processing. It has dNTPase activity against all standard nucleotides as well as RNA 5 trisphosphatase activity, which can play a role in the first stage of formation of the 5 cap structure of viral mRNAs. In MHV, nsp14 greatly enhances replication, for the proliferation and stability of the unusually large CoV genome. The recombinant viruses had a growth defect with mutations in the active site nsp14, and RNA synthesis had 15 times more mutations than wild-type viruses. Therefore, Nsp14 appears to play a role in preventing error or repairing the nucleotide composition during RNA synthesis. The recombinant HCoV-229E containing nsp14 active mutation has a severe RNA synthesis defect and no live virus can be recovered. In addition to reduced genome proliferation, specific defects in RNA synthesis were observed, such as inappropriate sizes of specific RNAs and changes in molecular ratios between individuals' RNA species. Nsp14 interacts with nsp10 and nsp16 to form a viral capitulation complex. Y2H and co-immunoprecipitation investigations show that nsp1 interplay with E, and sars3a44. The role of nsp16 methylation was explored in the viral duplication period and receive, that O-methylation acts as a diagnosis marker that aids the host cell to recognize its own RNA species, and react to imperfectly methylated cap structures. Nsp16 makes duplication feasible by camoflauging newly synthesized viral RNA be similar to host mRNA, thus blocks the induction of an interferon answer. This proposes that drugs that act on nsp16 cold interfere with viral replication both at the stage of prohibition trend of duplication, and in developing intracellular diagnosis and reflection to viral RNA species2.
In secondary lymph nodes, macrophages are the main cell of MHV, and recent studies show that cDCs can also be easily infected with MHV A59 or MHV JHM45. It is important to note that uncontrolled infection of cDCs by MHV is detrimental to the initiation of an adaptive antiviral immune response. In addition, laboratory studies have shown that neither macrophages nor monocyte-derived DCs respond to SARS-CoV infection with significant alpha IFN production. In contrast to the well-defined defense mechanisms of the adaptive immune response to MHV, the inherent immune response is not fully defined. Similarly, the importance of intrinsic safety mechanisms resulting from SARS-CoV remains unclear. The results showed that PBMC did not produce large amounts of cytokines and chemokines in air from SARS-CoV-infected individuals. The lack of a significant IFN response in PBMCs in patients with SARS-CoV may be due to partial inhibition of IFN type I signaling not only in nonlymphoid cells but also in pDCs46. Therefore, the potential of various non-structural SARS-CoV proteins that may inhibit or modulate type IFN responses in pDCs and other important target cells should be considered in future studies5.
CNS pathogens of chronic and viral rodents associated with myelin loss include two RNA virus models with good characteristics: TMEV, a member of the underdeveloped Picornaviridae, and MHV, a member of the Coronaviridae. Although CD8 + T cells are important in controlling the acute phase of both infections, these viruses can escape immune surveillance and cause chronic CNS infection by losing continuous myelin. Compatibility with nonCEACAM-1 tolerance cells can be expressed in collaboration with infected cells, prone to CEACAM-1 susceptibility, as well as independent CEACAM-1 infection in laboratory conditions. MHV initiates intracellular infection by the interaction of the S protein virus with its cellular receptor, the CEACAM-1 molecule. Analysis of S genes by MHV strains that show the selection of diverse pathogenesis of S gene mutation viruses and recombinant viruses with modified S genes all confirms that S protein is the main determinant of cell tropism and pathogenesis. But recombinant MHV analysis shows that in the absence of a dominant CD8 + T cell epithelium, other viral genes in addition to S genes also affect pathogenesis.
Various Pathogens of the Central Nervous System MHV
There are several different types of MHV, basic tropical regions and pathogenesis generally reflects the main serotype: MHV-1, MHV-2, MHV-3 and MHV-A59 are generally hepatic. John Howard Mueller (JHMV) and MHV-4 strains are usually nervous. The pathogenicity of MHV pathogenesis has long been apparent based on a variety of plaque sizes. The most accurate genetic and biological information is available for the relatively neurotic JHMV and MHVA59 strain. Data are focused on S protein because JHMV types with S protein changes or deletions show different prevalence. Recombinations that differentiate protein S in the context of heterogeneous virus confirm that JHMV S protein donates viral nerve, although JHMV background genes ignore A59 S protein in liver determination. The removal of the dominant T8 epithelial cell CD8 from the S protein, a multifaceted feature, and less pathogenic species complicates the pathogenesis. In general, viral strains infect most neurons and quickly lead to fatal encephalitis. Less pathogenic strains are mainly tropical glial and lead to persistent CNS infection. Efforts to increase neural communication with immunity showed no association with mRNA expression of the TNF-α, CCL5, or CCL2 gene. However, lethal JHMV infection increases IL-6, IL-1α, IL-1β, IFN-β, CCL3, CCL4, and MIP-2 compared with MHV-A59 infection, which is characterized by induction of prominent IFN-γ mRNA47. Surprisingly, the induction of IL-6 is associated with JHMV background genes, not with S protein. In contrast, by inducing unique chemokines and cytokines associated with central nervous system infection by JHMV and MHV-A59, non-lethal JHMV strains that differ only in S protein do not make a significant difference except for immune response magnitude. JHMV / A59 recombinants, using a highly viral JHMV parent, provide insight into the role of S protein and other viral proteins in pathogens. Pre-packaged MMP-9 secretion by neutrophils, re-regulation of adhesion molecules on the endothelial CNI, and possibly IL-6 action help eliminate BBB integration, which facilitates further entry of more inflammatory cells into the infected CNS. MMP-3, MMP-9, and MMP-12 mRNAs decrease either at the peak of JHMV-induced inflammation, or rapidly thereafter, contribution as a primary role in the development of the CNS environment48. Although, with the exception of MMP-9, their role in innate inflammatory-cell trafficking and CNS pathology is unclear6.
Spike Protein
The use of corona recombinant viruses, including MHV, TGEV, and IBV, has shown that S is a major determinant of tropical and pathogenic regions. The placement of the A59 spike gene with the very neonatal isolated spike gene on the part of the JHM virus is very neurological. The high neurotransmitters granted by S JHM are associated with rapid spread through the CNS, which may occur independently of the CEACAM receptor and a large number of infected neurons. However, the resulting Chemical virus is not as viral as the parents' JHM, at least in part because it makes the T8 cell response much stronger. The mechanisms that underlie the difference in immune response in the brain with the proximal strains of A59 and JHM are attractive and not understood at all. Replace of moderate SV protein of MHV-A59 liver with nonhepatotropic S JHM in recombinant viruses with the ability to induce hepatitis. Therefore, for recombinant viruses with A59 background genes, the ability to induce hepatitis largely depends on the ability of S to mediate entry into liver cells. However, the result is somewhat different in bodies where JHM-derived background genes are derived. The substitution of the JHM spike with A59 spike results in a chimeric virus which causes minimal infection of the liver and compel hepatitis Poorer; thus, in the attendance of JHM precedent genes, the spike of the A59 strain cannot mediate efficient infection of the liver 7. Therefore, in the presence of JHM background genes, S59 strain S is not able to effectively infect the liver. The mechanism by which JHM background genes inhibit liver infection is interesting. In a similar spike replacement test performed with IBV, the ectodomain protein S strain of viral M41-CK strain was used to replace the relevant area in the apathogenic IBV biota genome. The resulting chemical virus shows the laboratory cellular tropism phenotype M41-CK. However, the virus remains pathogenic. Therefore, the S M41-CK is not sufficient to infect the Kimrik virus. Thus, the S protein determines the tropics and therefore the pathogenesis; however, the S alone is not the primary determinant of pathogenesis and as the data show, other genes are involved in pathogenic phenotypes. The isolate, called MHV-4 or MHVSD, has the longest MHV HVR among JHM Ss, and in the absence of a CEACAM receptor, it is able to induce cellular fusion into the cell and spread the virus. It is possible that this ability is also related to a difference between S1 and S2, making it easier to evoke conformational changes in S that lead to fusion, which in turn is at least partially responsible. Similarly, deletions, as well as single-site mutations, have been shown in the HVR region to be present in intrusive neural effects. Mutations in both RBD from S1 and hepad repetition ranges in S2 have been shown to affect pathogenesis. Mutations in the RBD are likely to affect the interaction between S and the host cell, and therefore can affect the entry and virulence of the virus, while relocation uses a combined mechanism to affect the tropics. Variety in the amino-terminal transport terminal in TGEV and IBV; poor PRCoV elimination in the amine section of terminal S1 compared with malignant TGEV. Replacement of amino acids only in RBD, S310G, is responsible for the neurodegeneration of a JHM isolate. In addition, a single Q159L aminoacid replacement in this region removes the ability of MHV-A59 to infect the liver while having no effect on neurovirulence. The seeing that an aminoacid replacement in the RBD can confer a total loss of tropism to the liver when no infection affect the brain, while using alike Carcinoembryonic Antigen-Cell Adhesion Molecule (CEACAM) receptor, proposes that the surface of other molecule cells may does as cofactors or coreceptors in an organ specific method. An E1035D substitution within (Heptad repeats) HR1 may dominate the Q159L mutation, since a spike with both of these replacements confers hepatotropism on a recombinant MHV-A5949. For reinforcement of the(Receptor Binding Domain) RBD may cooperate with regions of the HR, escape mutants was chosen by resistance to a mon-oclonal antibody mapping to the receiver binding domain of S1 had point mutations in the domain of HR2, proposing a contact among these 2 physically gap portions of the spike50. In addition mutations within S1 may althogh affect host range; 21 amino-acid replacements and a 7-aminoacid insertion within the N-terminal domain of spike, but downstream of the RBD, allow MHV infection of the usually resistant hamster, feline, and monkey cells51. It is believed that SARS-CoV have jumped to humans from civets7. The conformity of SARS-CoV to humans perhaps implicates changes in the RBD. In matching of the spike protein of civets and humans, there are 6 aminoacid differences within the RBD of the spike. The spike protein of civet SARS-CoV has low dependence for the human ACE2 SARS-CoV receptor. Replacement of 2 amino-acids with the RBD of the human spike protein with those of the civet spike (N479K/T487S) approximately abrogates being able to infect (by using the single-round infection test) human cells presenting the SARS-CoV receptor7. Therefore, it is possible that the amino acids 479 and 487 are available for interactive and therefore specific species in the city, and the selection of viruses by replacing these residues makes it possible to adapt SARS-CoV to humans. Mutations in the S MHV safety epithelium have been reported as a mechanism for escaping the immune response and achieving viral stability. However, when the same inactive mutation entered a recombinant virus, depending on the genetic type of the virus and the strain of the infected mouse, the resulting virus varied from low to high in mice. Under similar circumstances, disabling the mutation in an external CD8 cell epithelium (gp33 of the lymphocytic choriomeningitis virus), which has been introduced to recombinant MHVs in an unnecessary gene, is easily attenuated in mice that have already been immunized against this epithelium and were selected. Therefore, the likelihood of an epithelium escaping depends on several factors, including the location of the epithelium in an essential protein versus an unnecessary protein and its effect on protein function, viral and age background genes, and mouse pressure. Epithelial T444 cell epitopes have been identified in S as well as M and N proteins from MHV and N proteins from TGEV and goat coronavirus. Studies with recombinant chimeric A59 / JHM viruses have shown that genes other than S play an important role in determining tropical regions. In fact, JHM genes destroy the ability of the A59 spike expression virus to cause hepatitis, and this is not due to replication but the genes at the '3 end of the genome. In addition, the rate of T cell response to recombinant MHVs and the possibility of viral clearance to occur, not diagnosis by the spike gene, but rather by context genes, re-encoded in the 3' end of gene. Therefore, other viral structural genes are clearly affected by pathogenicity. The SARS-CoV spike protein may play a role in pathogenesis by inducing lung interleukin-8 by activating MAPK and AP-1. Such activity was plotted to amino- acids 324 to 688 of the SARS-CoV spike. The activity was performed in epithelial cells and fibroblasts by using baculovirus presented SARS-CoV spike; the position of the sequencing cause of this function overlaps with the RBD, proposing that appendix to the ACE2 receiver might trigger this activation52.
Hemagglutinin-Esterase Protein
Coronavirus HE proteins have not received much attention in the past, probably because they are unnecessary for the proliferation of tissue culture or, in the case of MHV, viruses in mice. While HE is essential for proliferation in cell culture, it is unnecessary; it is necessary and sufficient for S entry. Therefore, the role of HE in coronavirus infection is still unclear and deserves further investigation. Observing that the expression of HE is unnecessary for the viral life cycle indicates that HE may be involved in animal defects. HE has long been thought to be involved in an acute or chronic disease caused by MHV, possibly as a determinant of cellular tropism, or may be amplified by attachment or proliferation. Help the virus. There are both initial data that agree and disagree with this hypothesis. Compared to a type that has lower HE expression, a higher level of mortality as well as an increase in neuronal infection was associated with a type of JHM that indicates a high level of HE. However, contrary to these studies, Lee et al. Reported that in mice infected with JHM, defective HE viruses accumulated in the brain and spinal cord. These studies were performed before the inverse genetics were available for MHV, so they were unable to distinguish the effects of HE and the effect of other genes on the comparison of MHV isolates. A recent study of the pathogenesis of recombinant isogenic recombinant viruses expressed the expression of the wild type of HE protein, in which the activity of acetylsalicylic acid was eliminated by mutation and did not indicate HE polypeptide. Surprisingly, both viruses, which represent HE polypeptides, were more viral in rats when injected into mice. This result could be in line with the model in which HE may enhance the attachment of the virus, and the scattering of enzymes containing tosialic acid may indicate that the binding range of the cialic acid is separate from the ester field. Our hypothesis is that for MHV infection from the CNS, it is the HE binding activity that increases the virus in certain cell types, and at least in the CNS, esterification activity is not important for advanced development. Stress activity may be more important in other organs, such as the respiratory tract, where the virus may need to pass through the mucosa or be able to separate cells that may not be productively infected, both of which appear to be. Normaminidases may work. In the case of influenza virus, it has recently been shown that norepinephrine, for a specific type of cialic acid, determines the amount of cells disinfected with respiratory extract and the outcome of the pathogen. Thus, comparatively, HE MHH may also play a role in tropical regions.
Membrane Protein
The M protein is the most abundant protein of virion membrane. Aside from its role in viral assembly, it is believed that the coronavirus M protein have functions in host interactions. It might be O glycosylated (groups I and III) or N glycosylated (group II). During the time that glycosylation is not fundamental for viral assembly or infectivity, the glycosylation state of M protein perhaps has a role in virus-host interaction. For Transmissible Gastroenteritis Virus (TGEV), the M protein has been shown to have interferogenic activity, and mutations in the M protein ectodomain that impair N glycosylation decrease this activity. For MHV, the selection of recombinant viruses with N, O, or no glycosylation demonstrated that while the glycosylation state of M protein does alter the ability to replicate in vitro, it may affect the ability to induce IFN-_ in vitro and also to replicate in the liver in vivo53.
Nucleocapsid Protein
Furthermore, the structural N protein has a role in transcription and also in pathogenesis. Expression of N protein is essential for the effective recovery of virus from infectious cDNA clones and recently has been shown to enhance the proliferation of HCoV-229E genome RNA54. MHV protein N is involved in hepatitis flumint. Infection of mice with MHV-3 pressure strongly stimulates the hepatic expression of the fgl2 gene, which expresses a novel protein protein molecule, a bronchogenic protein such as 2, which is encoded in the liver and endothelial cells of the liver. The ability to regenerate the transcription of these gene maps into the nucleococcid gene is associated with the progression of hepatitis fulminan. While MHV proteins are generally restricted to the cytoplasm, nucleolus proteins from coronaviruses representing groups I, II, and III have been shown to be localized to the nucleus as well as the cytoplasm. The report suggests that protein N delays or arrests the cell cycle, most likely in the G2 / M stage, possibly inhibiting cytokines.
Small Envelope Protein
Along with M and E proteins, it plays an important role in viral assembly. Protein E when expressed alone or in combination with M, forms particles such as viruses, surprisingly, it is possible to select a recombinant MHV by removing the E gene. Such a recombinant MHV has low infection and poorly replicates, indicating that while it is unnecessary for MHV, E plays an important role in the production of infectious virus. The recently presentation indicate that the E protein of SARS-CoV has cation selective ion channel activity. Whenever the task of this activity is still unknown, the E protein ion channel can act at the budding site to enhance viral morphogenesis and assembly. E protein‘s other task is in the hostvirus interaction, especially in induction of apoptosis. E induces apoptosis in vitro in MHV-A59-infected 17Cl-1 cell via a caspase-dependent mechanism; such apoptosis is suppressed with high level of Bcl-2 expression55. Inhibition of MHV-induced apoptosis causes virus production in late infection and suggests that apoptosis may be a host response that limits the level of virus production. It is unknown at this time what he will do after leaving the hospital. Similarly, the SARS-CoV protein E has been shown to induce apoptosis when expressed in T Jurkat cells, and this activity is inhibited by the expression of the Bcl-xL antibody protein. The authors said that T cell apoptosis may contribute to SARS-CoV lymphopenia, which is seen in most SARS patients.
Replicase Proteins
Identification proteins can affect tropical regions and pathogenesis by determining the rate of viral replication, perhaps through interactions consisting of, 5 or, 3 UTR genomes as a result of virus transmission, with cell type factors, or with immune response elements. Several enzymatic activities that are expected to be encoded in ORFs 1a and 1b, as described above, can play a role in disrupting various aspects of host metabolism cells. Studies of MHVs chimeric A59 / JHM, in which the A59 proliferation gene is expressed with JHM structural genes and vice versa, show that this proliferation is not a significant determinant of the difference in tropical and pathogenic differences between the two strains (severe encephalitis versus Hepatitis) These are the three parts of the genome that are responsible for the pathogenic properties. This is in line with the MHV's ability to induce hepatitis maps into the nucleococcal gene.
Group-Specific Proteins
While there are no specific examples of coronavirus proteins involved in defending the host's innate immune system in defense against antibodies, it is quite possible that one or more unnecessary coronavirus proteins, such as "side effects," can be infected during infection. Act inside the body. The coronavirus genes that encode these proteins are sometimes referred to as smallORF genes because they protect against each group of coronaviruses. There are several examples of recombinant viruses in which deleting the expression of ORFs may alter the pathogenic phenotype. In support of a role in the pathogenesis for ORF 2a-encoded 30-kDa protein is the report of a recombinant MHV in which a mutation in ORF 2a is associated with reduction in animals56. ORF 2a encodes phosphodiesterase cycles. It is interesting to speculate that such an action may jeopardize the host's response. It has been suggested that cyclic phosphodiesterase, along with the predicted activity of ADP-ribose1 -phosphatase, encoded by ORF1a, may have successive steps in the processing of intermediate tRNA synthetic products. In the case of TGEV pork coronavirus, gene expression 7 is not necessary for reproduction. However, a recombinant virus in which the expression of gene 7 has been ruled out reduces the rate of virus replication in the lungs and intestines, indicating that the product of gene 7 has an effect on reproduction within the body. ORFs 3a and 3b TGEV are also not required for proliferation. In addition, recombination by eliminating these ORFs indicates a wild-type stability that is effectively propagated in animals and shows only a very small reduction. The human SARS-CoV genome codes several small open reading fames (ORFs 3a, 3b, 6, 7a,7b, 8a, 8b, and 9b) that are presumed to encode eight group-specific accessory proteins57. ORFs 8a and 8b may be caused by the removal of animal SARS-CoV isolates in ORF 8. Therefore, ORF 8 may not be necessary for proliferation in humans. Two types of these ORFs have been reported during infection, and some coded proteins have been identified. ORF7a-encoding protein The ORF7a amino acid protein contains a sequence of spiral and C-terminal signal signals, most likely an I-type membrane protein. ORF 7a protein is expressed in Vero E6 cells and patients' lungs, but not in viruses. ORF 3a protein can interact with structural proteins M, E, and S, as well as with ORF 7a protein. In addition to protein E and protein N, ORF 7a, ORF 3a and ORF 3b have been reported to cause apoptosis. Because all of these studies involve overexpression of individual proteins, it is difficult to determine what role they may play during infection. It will be important to analyze recombinant viruses with specific mutations that clearly indicate the role of each specific protein in apoptosis. Recent data suggest that these small ORFs may be different between SARS isolates. For example, there are reports that in the SARS-CoV isolates from humans, there are deletions in an ORF 8 that lead to two types of ORF, 8a and 8b. The difference in S-gene sequences was the most significant difference observed between the SARS-CoV animal genome and the animal genome. In another study, removal of 45 nucleotide "within the frame", occurred in ORF 7b after three SARS-CoV crossings in tissue culture. Elimination was observed in ORFs 6 to 8 in SARSCoV compatibility with primary cell culture. While it is tempting that one or more of these ORFs participate in adapting to the human host or overthrowing the host's innate and immune responses, it involves issues related to discrimination and exchange of views.
SARS Pathogenesis
A SARS disease model is presented, which includes three stages: viral proliferation, hyperactivity in the immune system, and pulmonary destruction. Lung SARS pathology has been associated with alveolar damage, proliferation of epithelial cells, and increased macrophages. A large number of giant cells in different parts of the macrophage or epithelial origin are considered to be similar to sinusitis, which is characteristic of many coronavirus infections. The lymphopenia, hemophagocytosis in the lungs, and white-pulpatrophy of the spleen observed in SARS patients are reminiscent of those reported for fatal flu virus subtype H5N1 disease in 1997. Interestingly, the presence of hemophagocytosis supports a cytokine deregulation58. Recent events have been based on autopsies of SARS patients, in which SARS is a systemic disease with widespread extrapulmonary spread and leads to viral shedding in respiratory secretions, feces, urine and even sweat. Cytokines released by stimulated macrophages in alveoli may play a role in SARS pathogenesis. SARS-CoV infection of macrophages in laboratory conditions leads to protein detection and protein virus use, but no virus particles are produced. In opposite to the case for flu A virus and HCoV-229E, no IFNa/b react is diagnosed in macrophages, despite the induced explain of chemokines for example CXCL10/IFN-γ-inducible protein 10 and CCL2/monocyte chemotactic protein 159. Interestingly, SARS-CoV has recently been reported to proliferate in SARS peripheral blood peripheral mononuclear cells. Regunathan et al. PBMC gene expression analysis of 10 SARS patients was analyzed in comparison with healthy groups. PBMC gene expression analysis of SARS patients using a microarray platform containing more than 8000 gene sequences shows that the response of SARS patients appears to be more than an innate inflammatory response rather than a specific immune answer against a viral infection. Reghunathan et al60 do not have a significant effect on class I complex genes or major cytokines, including IFNs or genes involved in complementary cytolism, in terms of significant compatibility with complex class I cells or important cytokines. They concluded that the immune response to SARS-CoV may be different from other viral infections, or that the virus may be inflexible and deadly using an unusual strategic operating system. Lymphopenia and increased viral load in the first 10 days of SARS indicate the escape of the immune system by SARS-CoV. The use of SARS-CoV-infected cells in response to beta-IFN receptors has not been reported using dendritic cells derived from primary human myeloid and epithelial cell line 293. Love et al. proposed the mechanism of escape from the immune system by SARS-CoV in DC. Spiegel et al. showed that SARS-CoV inhibited interferon-mediated growth by inhibiting alpha IFN induction by interfering with activation of the 3 IFN regulatory factors. The mechanism of lymphopenia is unknown. Rapid reduction of CD4 and CD8 cells may be associated with adverse outcomes. Due to the lack of ACE2 expression in T lymphocytes and B cells, it is unlikely that SARS-CoV lymphopenia is caused by a direct viral infection. Instead, it has been suggested that apoptosis of uninfected lymphocytes may be indirectly maintained, especially in patients with SARS. In this sense, various SARS-CoV proteins have been suggested to stimulate the enzyme apoptosis. These include ORF7a, ORF 3a and ORF 3b in addition to protein E and protein N. Inhumane pheromones, domestic cats, mice, Syrian mice and golden hamsters have been experimentally infected with SARS-CoV. These animals support the recurrence of acute virus in the upper and lower respiratory tract, although there is a difference in results. In contrast, pigs and chickens can be experimentally infected with SARS-CoV, but these species do not support effective SARS-CoV proliferation7. (Figure 2)
Figure 2.Some symptoms of COVID-19 https://upload.wikimedia.org/wikipedia/commons/3/33/Symptoms_of_coronavirus_disease_2019_4.0.svg
Clinical Symptoms of COVID-19
Huang et al, perfomrd a study about Clinical symptoms of COVID-19, Findings By Jan 2, 2020, from 41 patients that admitted hospital had been identified as having laboratory-confirmed 2019-nCoV infection. 73% of the infected patients were men (30 of 41); 32% had underlying diseases 13 patient, 20% diabetes 8 patient, 15% hypertension 6patient, 15% cardiovascular disease 6 patient. Median age was 49·0 years (41·0–58·0). 66% patients 27 of 41 had been exposed to Huanan seafood market. One family cluster was found. Common symptoms at onset of illness 98% were fever (40 of 41 patients), 76%cough (31 patients), and 44% myalgia or fatigue (18 patients); 28% common symptoms were sputum production (11 of 39 patients), 8%headache (3of 38 patients), 5% haemoptysis (2 of 39 patients), and 3% diarrhoea (1 of 38 patients). 55% Dyspnoea developed in 22 of 40 patients. Median time from illness onset to dyspnea was 8·0 days (5·0–13·0)). 63% had lymphopenia 26 of 41 patients. All 41 patients had pneumonia with abnormal findings on chest CT. Complications included 29% acute respiratory distress syndrome (12 patients), 15% RNAaemia (6 patients), 12% acute cardiac injury (5 patients) and 10% secondary infection (4 patients). 13patients were admitted to an ICU (32%) and six patients died (15%). Compared with non-ICU patients, ICU patients had higher plasma levels of IL2, IL7, IL10, GSCF, IP10, MCP1, MIP1A, and TNFα 61.
Demographic and Clinical Characteristics
Of all 1,324 patients recruited as of January 29th, 222 had a suspected diagnosis (16.8%) and were therefore excluded. The core data sets (including clinical outcomes and symptoms) of 3 patients were lacking due to the incompleteness of original reports, hence this report delineates 1,099 patients with 2019-nCoV acute respiratory disease (ARD) from 552 hospitals in 31 provinces-level municipalities. The demographic and clinical characteristics indicated that 2.09% were healthcare workers. A history of contact with wildlife, recent travel to Wuhan, and contact with people from Wuhan was documented in 1.18%, 31.30% and 71.80% of patients, respectively. 483 patients(43.95%) were local residents of Wuhan. 26.0% of patients outside of Wuhan did not have a recent travel to Wuhan or contact with people from Wuhan. The median incubation period was 3.0 days (range, 0 to 24.0). The median age was 47.0 years (IQR, 35.0 to 58.0), and 41.9% were females. 2019-nCoV ARD was diagnosed throughout the whole spectrum of age. 0.9% of patients were aged below 15 years. Fever (87.9%) and cough (67.7%) were the most common symptoms, whereas diarrhea (3.7%) and vomiting (5.0%) were rare. 25.2% of patients had at least one underlying disorder (i.e., hypertension, chronic obstructive pulmonary disease(COPD)). On admission, 926 and 173 patients were categorized into non-severe and severe subgroups, respectively. The age differed significantly between the two groups (mean difference, 7.0, 95%CI, 4.4 to 9.6). Moreover, any underlying disorder was significantly more common in severe cases as compared with non-severe cases (38.2% vs. 22.5%, P<0.05). There were, however, no marked differences in the exposure history between the two groups (all P>0.05) 62.
Radiologic and Laboratory Findings at Presentation
Of 840 patients who underwent chest computed tomography on admission, 76.4% manifested as pneumonia. The most common patterns on chest computed tomography were ground-glass opacity (50.0%) and bilateral patchy shadowing (46.0%). The representative radiologic findings of two patients with non-severe 2019-nCoV ARD and another two patients with severe 2019-nCoV ARD. Despite these predominant manifestations, 221 out of 926 in severe cases (23.87%) compared with 9 out of 173 non-severe cases (5.20%) who had no abnormal radiological findings were diagnosed by symptoms plus RT-PCR positive findings(P<0.001). Severe cases yielded more prominent radiologic abnormalities on chest X-ray and computed tomography than non-severe cases (all P<0.05). On admission, 82.1% and 36.2% of patients had lymphopenia and thrombocytopenia, respectively. Overall, leukopenia was observed in 33.7% of patients. Most patients demonstrated elevated levels of C-reactive protein, but elevated levels of alanine aminotransferase, aspartate aminotransferase, creatine kinase and D-dimer were less common. Severe cases had more prominent laboratory abnormalities (i.e., leukopenia, lymphopenia, thrombocytopenia, elevated C-reactive protein levels) as compared with non-severe cases (all P<0.05) 62.
Treatment and Complications
Overall, oxygen therapy, mechanical ventilation, intravenous antibiotics and oseltamivir therapy were initiated in 38.0%, 6.1%, 57.5% and 35.8% of patients, respectively. All these therapies were initiated in significantly higher percentages of severe cases (all P<0.05). Significantly more severe cases received mechanical ventilation (non-invasive: 32.37% vs. 0%, P<0.001; invasive: 13.87% vs. 0%, P<0.001) as compared with non-severe cases. Systemic corticosteroid was given to 18.6% of cases and more so in the severe group than the non-severe patients (44.5% vs 13.7%, p<0.001). Moreover, extracorporeal membrane oxygenation was adopted in 5 severe cases but none in non-severe cases (P<0.001). During hospital admission, the most common complication was pneumonia (79.1%), followed by ARDS (3.37%) and shock (1.00%). Severe cases yielded significantly higher rates of any complication as compared with non-severe cases (94.8% vs. 72.2%, P<0.001).
Clinical Outcomes
The percentages of patients being admitted to the ICU were 5.00%, requiring invasive ventilation were 2.18% and 1.36%were death. This corresponded to 67 of patients (6.10%) having reached to the composite endpoint.Results of the univariate competing risk model indicated that severe pneumonia cases (SDHR, 9.803; 95%CI, 4.06 to 23.67), leukocyte count greater than 4,000/mm3 (SDHR, 4.01; 95%CI, 1.53 to 10.55) and interstitial abnormality on chest X-ray (SDHR, 4.31; 95%CI, 1.73 to 10.75) were associated with the composite endpoint62.
Diagnosis of COVID-19
The diagnosis of coronavirus is unnecessary in most cases of infection because most patients have mild or moderate syndrome with a strong prognosis. However, it may be necessary to identify an etiological factor in epidemiological studies, especially during epidemic outbreaks. Since the new Coronavirus-2019 has not been found in humans before, no specific vaccine or treatment has been provided. In the current state of emergency, the number of cases is rapidly increasing. Therefore, it is very important to diagnose all suspected cases as soon as possible and to remove them quickly, to cut off the source of the infection. Conventional diagnostic testing methods, such as assessment to detect antiviral antibodies or viral antigens, have been clinically developed and used. New diagnostic solutions, including RT-PCR and microscopic-based measurements, may be effective in monitoring epidemiological measures, along with preventive measures. Coronavirus Novin-2019 nucleic acids can be detected in samples such as nasopharyngeal swabs, sputum, lower respiratory tract secretions, blood, and feces19.
Ways to Transmit COVID-19
Novel coronavirus-2019 is thought to be transmitted through droplets, close contact, aerosols, and possibly fecal-oral transmission, and patients in the incubation period can transmit the virus to others. The virus receptor distribution can explain pathogenic mechanisms, clinical manifestations, and routes of transmission of the Novel Coronavirus-2019. The enzyme converting angiotensin 2 (ACE2) has been suggested as a receptor for the Novel coronavirus-2019, which is essential for its entry. Comprehensive expression of ACE2 in various cells, such as lung AT2 cells, esophageal sphincter, epithelial cells, and absorption enterocytes of the ileum and large intestine, may play a role in multinodular coronavirus infection. The mouth is a potential route for a Novel 2019 coronavirus infection19. Although the route of transmission has not been clearly identified, airborne droplets from infected patients may be the main route of transmission or blood flow through the mouth and teeth is a route for transmission in one case of the indicator7. So far, the exact answer to the question "Whether animals are involved in the transmission of this disease has not been determined exactly, but there is a high probability for it, given the history of contact with camels and some animals in these patients and the genetic similarity of the new virus with Coronavirus." Bat, animals are likely to be the reservoir of the disease, and people have contracted the disease after contact with them, however, the existence of an animal reservoir for this disease has not yet been determined8. It shows the fecal-oral route as a possible transmission route for chickens9.
The Reasons for the Spread of the Virus
Due to the intensive trade and uncontrolled movement of poultry and people between the provinces of Iran, the distribution of Gammacoronavirus in poultry farms is very possible9. Kittens are usually infected between the ages of 6 and 8 weeks, while maternal-derived antibodies are eliminated by contact with the feces of mothers or other FCoV-excreted cats10. In addition, many proteins are aquatic organisms that have never left the environment in which the forms of prokaryotic and eukaryotic life have evolved along with the corresponding viruses. The primary environment also provides protection against ultraviolet radiation and ultraviolet radiation, providing a suitable means of dispersing the virus through diffusion, as well as keeping the virus alive11.
Ways to Prevent and Control COVID-19
As no vaccine has been successfully developed to prevent COV-19 until March 1, 2020, public health measures to reduce infection are essential to reduce the global spread of the virus in order to reduce the harm associated with COV-19. The experience of the early stages of pneumonia of the Novel Coronavirus-2019 strongly emphasizes that the history of travel (especially from China), instead of chest radiography, is of great importance for early detection and isolation of cases of Coronavirus Novin-2019 pneumonia. Restricting travel and human-to-human transmission in order to reduce secondary infections in close contact with health care personnel and prevent further spread of the disease from China is essential. Based on previous experience in the management of SARS and MERS infections, the World Health Organization (WHO) has intervened to control infection to reduce the overall risk of transmitting acute respiratory infections, including avoiding close and direct contact with people with acute respiratory infections, frequent hand washing, especially after Direct contact with sick people in their environment and avoidance of unprotected contact with domestic and wild animals is strongly recommended. In addition, people who have symptoms of an acute respiratory infection should follow the coughing etiquette, which is to keep a distance, cover up coughing and sneezing with a disposable towel or cloth, and wash your hands frequently. In health care centers, increasing standards of infection prevention and control in hospitals has been recommended, especially in the emergency and infectious wards. It is also possible that reducing stress and anxiety in patients to prevent suppression of the immune system can be effective in controlling COVID-19 infections. The US Centers for Disease Control and Prevention (CDC) has provided temporary clinical guidelines for the prevalence of COVID-19 to implement offensive measures to slow the transmission of the Novel Coronavirus-2019 in the United States. These measures include identifying cases in the United States, as well as assessing and taking proper care of travelers coming to the United States from the mainland, especially China. All efforts have been made to slow down the spread of the disease in order to provide or buy time to better provide public health care systems, to better describe COVID-19 to guide public health recommendations and to develop timely diagnosis, treatment and vaccination19. Three types of data were searched, including historical historical records, human research evidence, and current prevention programs.
First Classical Historical Records
Records for the prevention of epidemics were sought in ancient CM books, including history, principles of treatment, medicine, and the use of CM to prevent epidemics.
Second, Human Research Studies
They have been conducted to evaluate the preventive effects of CM on infectious respiratory viral diseases. The entry criteria are as follows. Study design: clinical trial, cohort studies and other uncontrolled population studies. Population: Population at risk for SARS or H1N1 flu. Intervention: Oral CHM formulas, including decoctions, granules or patents. Control: placebo, empty or without control group. The result was that the rate of infection was confirmed as a disease test.
Third Current Prevention Programs
CM Prevention Programs for COVID-19 issued by the state or provincial health authorities in China. Due to the fact that some provinces have regularly updated their programs based on local prevalence and clinical performance, the latest versions of the programs for analysis are included in this study. CHM formula to prevent pestilence in ancient CM: Pestilence prevention and treatment theory written about 2000 years ago. Healthy qi in the body using preventive medications (Xiaojin Dan in Huangdi's Classic, the first recommended CM formula to prevent pestilence), healthy diet, exercise, etc., to resist the invasion of foreign pathogens and more. These two principles of epidemiological prevention have so far been pursued by specialist physicians in CM. In his book, the famous physician SUN Si-miao explained the basic prescriptions, which are worth thousands of gold for emergencies, the basis of drugs to prevent pestilence: pestilence comes from nature, so to prevent it, we must prepare medicinal plants. Let's say that they also originate from nature. If people know and take preventative medicine, people will not be infected.
Evidence for Using the CHM Formula to Prevent SARS
Three studies, including 1 controlled study and 2 cohort studies performed during the SARS epidemic were identified. To evaluate a plant formula for preventing SARS (without plant interventions in the control group), Love et al. Conducted a control study and conducted it in Hong Kong, China. The sample size was 16,437 people (1,063 in the plant group and 15374 in the non-plant group) and all participants in this hospital were hospital care staff including doctors, nurses and other employees. The results showed that none of the participants who received the modified formula of Yupingfeng powder with Sangju decoction were infected with SARS, while 64 of the 15347 individuals (0.4%) in the non-herbal group were infected with SARS. Nineteen cases (1.8) showed minor side effects after 14 days of taking herbal medicines, including diarrhea, sore throat, dizziness, and nausea. Both single cohort studies were performed in Beijing, China with sample sizes of 3561 and 163, respectively. All participants were medical staff from two hospitals, where SARS patients were recruited and treated during the study period. The courses of herbal formulas for prevention were 6 days and 12-25 days, respectively. The formula used in these studies was both the classic formula of Yupingfeng powder with some herbs to eliminate fever and detoxification. The results showed that none of the participants who were taking the herbal medicine were infected with SARS in the two studies.
Evidence from the CHM Formula for Preventing H1N1 Flu
Four studies were identified, including 3 randomized controlled trials and 1 uncontrolled randomized clinical trial. All studies were conducted during the outbreak of influenza H1N1 in mainland China and published in Chinese. In these studies, participants were exposed to high-risk environments such as hospitals and schools where the H1N1 flu occurred. The total sample size was 25636 individuals with the largest 25329. And 3 blank control items were used. The course of herbal formulas was from 3 to 7 days, while the follow-up was 5 to 30 days. The measurement of the result was the rate of H1N1 flu infection detected by laboratory serology. One study reported no side effects while others did not. Data on influenza H1N1 infection were collected from 4 studies in meta-analysis. The results showed that the degree of contamination in the group of herbal formulas was significantly lower than the control group (relative risk 0.36, confidence interval 95 ٪ 0.24-0.52), sensitivity analysis was performed to remove non-RCT and similar results (0.36 RR, 95% CI 0.21-0.62, P <0.01) showed that COVID-19, as an emerging acute infectious respiratory disease, lacks effective methods for controlling and treating infection and makes sense of strategy. Effective Interventions Use Traditional Medicine Intervention to Prevent It. Based on a comprehensive analysis of prevention programs issued by 23 provinces since the outbreak of COVID-19, we found that the basic principles of CM in Jal of COVID-19, strengthened to protect and provide defense against foreign pathogens. In addition, the characteristics formula CHM to prevent pestilence in ancient times and was similar to SARS in 2003. Six Consumer Plants: Astragali Radix (Huangqi), Glycyrrhizae Radix Et Rhizoma (Gancao), Saposhnikoviae Radix (Fangfeng), Atractylodis Macrocephalae Rhizoma (Baizhu), Lonicerae Japonicae Flos (Jinyinhua) Astragali Radix (Huangqi), Saposhnikoviae Radix (Fangfeng) and Atractylodis Macrocephalae Rhizoma (Baizhu) are all compounds of a classic herbal formula of Yupingfeng powder, to strengthen the protection against external pathogens. In a controlled study by Laos et al. of the CM formula to prevent SARS, feng yuppung powder was also a major ingredient. Some studies have confirmed that funging yuping powder has antiviral, anti-inflammatory and immune system effects. Japonicae Flos (Jinyinhua) and Forsythiae Fructus (Lianqiao) are the main components of Yinqiao powder, a classic formula used to prevent and treat respiratory infectious diseases in ancient times. An experimental study showed that the effect of Yinqiao powder on the prevention and treatment of upper respiratory tract infections can be explained by its antibacterial and antiviral properties and improve the function of the mucosal immune system. A large-scale randomized trial, Showed that Yinqiao powder plus the heat purification formula can reduce the time to cure fever in patients with H1N1 in influenza virus infection. China has not issued a plan to prevent CM for COVID-19. The reasons may, in the first place, be based on CM's theory of treatment due to differences in individual, regional, and seasonal factors in the incidence and distribution of disease, and second, the lack of strong evidence from the CM formula for COVID-19. By comparing and analyzing prevention programs issued by the provincial level, we also found that there is little logical difference in the recommended herbal formulas and prescribing principles. For example, due to the dry climate in northern China, there are one or two additional plants in the formula, such as Glehniae Radix and Ophiopogonis Radix, while in the south due to the humid climate of fragrant plants with moisture removal function and Turbidity is used in formulas such as Pogostemonis Herba (Huoxiang) and Eupatorii Herba (Peilan). Individual differences in prevention programs were also considered in some provinces. There are two or more formulas recommended in 18 provincial programs that apply to different populations, including the elderly, children, pregnant women, or patients with chronic disease, population correlation in close counteract with patients with COVID-19, and so on. . In addition, the 7 provinces or municipalities of the province (Beijing, Tianjin, Shanxi, Henan, Hunan, Shandong, and Yunnan) recommend formulas according to the types of CM body compounds. We recommend that you consider safety when using the CHM formula to avoid COVID-19, especially when used for a long time. It should also be noted that no decoction prevention recommendations have been reported in the 12 Provinces Program. According to the plans of other provinces, it is appropriate to do the decoction for 1 week. We do not recommend that everyone take CHM to prevent COVID-19. The results showed that these formulas contained 54 different plants, of which 19 plants were used for 3 times or more in preventive formulas for the general population of Radix astragali (Huangqi) and Glycyrrhizae Radix ET Rhizoma (Gancao). For future studies, we recommend prospective cohort studies, RCTs, or registry studies to assess the impact of CHM formulas on COVID-19 prevention. Currently, since COVID-19 is not yet controlled, we expect a series of prospective demographic studies to begin with accurate design and large sample, protocol registration, ethical verification, and timely implementation to provide credible evidence. Therefore, based on historical records and clinical evidence of SARS and influenza H1N1, the CHM formula could be an alternative method of preventing COVID-19 in high-risk individuals while awaiting successful vaccine production. Futuristic demographic studies are designed to assess the effects of CM prevention3.
Vaccine Strategy Against SARS
Previous experiences with coronavirus vaccines are related to SARS vaccine progress. Several investigations’ aim at passive and active immunization have used the animal models for SARS duplication. Subbarao et al. 63 demonstrated that passive transmission of immune serum protects naive BALB/c mice from SARS-CoV infection. Various studies have shown that human monoclonal antibodies approve some protection opposite SARS. Traggiai et al.64 Have created an improved method for Epstein-Barr virus transformation of human B cells. This method was used to analyze the memory repertoire of a patient who recovered from SARS-CoV infection and to isolate monoclonal antibodies specific for different viral proteins. Although some of these monoclonal antibodies exhibited in vitro neutralizing activity, only one of such antibodies conferred protection in vivo in a mouse model of SARS-CoV infection. Human IgG monoclonal antibodies against SARS with in vitro neutralizing activity and protection in a ferret model have been found using phage display libraries65. In both mouse and ferret models, administration of human monoclonal antibody with in vitro neutralization function decreased SARS titers in the lungs(3- to 6-log10-unit decrease), also protecting from lung pathology in ferrets. Several investigations were directed about the creation active immunization strategies. Inactivated virions, recombinant antigen, DNA vaccines, adenoviral vectors, vaccinia virus Ankara and recombinant parainfluenza virus type 3 vectors, and rhabdovirus-based vectors are being studied. Inactivated SARS vaccines have been presented to extract systemic humoral immunity in mice and high titers of spike-specific antibodies that block receptor binding and virus entry in cell culture66. Furthermore, UV-inactivated virion induced regional lymph node T-cell proliferation and high levels of cytokine (IL-2, IL-4, IL-5, IFN-γ, and TNF-α) production upon restimulation with inactivated SARS-CoV virions in vitro. Althogh, none of these investigations have addressed whether inactivated total SARS-CoV virions confer protection from virus challenge. Zhou et al.67 Have presented that inactivated SARS-CoV induces humoral and mucosal immunity opposite compete with SARS-CoV in rhesus monkeys. SARS-CoV spike glycoprotein68, M69, and Nucleocapsid70 have been appraised as candidate vaccines, using DNA immunization in mice. Interestingly, DNA vaccination can induce humoral and cellular immunity opposite SARS-CoV in the mouse model. Yang et al.71 Explained that a DNA vaccine encoding the codonoptimized SARS spike glycoprotein induces neutralizing antibody like T-cell answers. Protection from SARS-CoV compete was mediated by a humoral immune response but not by a T-cell-dependent function. Zeng et al.72 Have introduced that mice immunized by plasmids encoding fragments of S1 developed a Th-1 antibody isotype switching. A DNA vaccine encoding calreticulin linked to the nucleocapsid creates strong Nspecific humoral and cellular immunity and protects mice opposite a vaccinia virus expressing nucleocapsid. A prime-boost admixture of DNA and whole killed SARSCoV vaccines elicited higher antibody responses than DNA or total killed virus vaccines alone73. Adenovirus-based vaccination strategies opposite SARSCoV, using duplication-defective adenovirus type 5 vectors explaining structural SARS proteins (S1, M, and N), have also been reported. Vaccinated rhesus macaques developed antibody reactions opposite fragment S1 of spike, virus-neutralizing antibody feedback, and T-cell reaction opposite the nucleocapsid74. Similarly, Zakhartchouk et al.75 Explained that vaccination of C57B/L6 mice with adenovirus type 5-expressing nucleocapsid extracted SARS-CoV-specific humoral and T-cell-mediated immune reaction in C57B/L6 mice. The highly attenuated modified vaccinia virus Ankara (MVA) has been used to express the spike glycoprotein of SARS-CoV in vaccination experiments using the mouse76 and the ferret77 models, with different results. Intranasal and intramuscular (IM) administration of MVA encoding the SARS-CoV spike protein led to the induction of a humoral immune reaction in BALB/c mice, the same reduced viral titers in the respiratory tract. In ferrets, vaccination with MVA encoding the spike or nucleocapsid induced a powerful antibody reaction; however, it did not prevent virus infection and spreading. Liver inflammation (in the absence of viral antigen) was found in all MVA-spike-vaccinated ferrets and in only one MVA-nucleocapsid-vaccinated animal after challenge with SARS-CoV78. Inflammation in the livers of ferrets vaccinated with MVA-nucleocapsid was the same as that in the MVA control group. It is important to note that this study did not find any clinical disease in ferrets after infection with SARSCoV (Against others79). These authors pointed out that although their results need to be further studied, they may proposed antibody-dependent enhancement, the same as the case for FIPV. For this feline coronavirus, antibodies acquired either through a passive transfer of immune serum against the spike protein of FIPV or by immunization with a recombinant vaccinia virus expressing the spike protein lead to accelerated infection by the antibody-dependent enhancement mechanism80, 81. Recombinant bovine-human parainfluenza virus type 3 vector (BHPIV3) is a version of bovine parainfluenza virus type 3 in that genes encoding the bovine parainfluenza virus type 3 major protective antigens, the fusion (F) and hemagglutinin- neuraminidase (HN) glycoproteins were replaced with their counterparts from human parainfluenza virus type 3. BHPIV3 is being developed as a live attenuated, intranasal pediatric vaccine against human parainfluenza virus type 3. Immunization of African green monkeys with a single dose of BHPIV3 expressing SARS-CoV spike protein administered via the respiratory tract induced the production of SARS-CoVneutralizing antibodies82. A recombinant BHPIV3 expressing SARS-CoV structural protein (S, M, and N) individually or in combination has been evaluated for immunogenicity and preventive efficacy in hamsters, which support both SARS-CoV and BHPIV3 duplication in lungs83. A single intranasal administration of BHPIV3 expressing the SARS-CoV spike protein induced a high titer of SARS-CoV-neutralizing antibodies, only twofold less than that induced by SARS-CoV infection. In the absence of spike, expression of M, N, or E did not induce a diagnisible serum SARS-CoV-neutralizing antibody reaction. Immunization with BHPIV3 expressing spike provided whole protection counteract SARS-CoV challenge in the lower respiratory tract and partial protection in the upper respiratory tract. Faber et al.84 Have created recombinant rabies virus expressing the spike or the nucleocapsid protein of SARSCoV. These vectors induced a neutralizing antibody reaction in mice. Those authors concluded that the use of rabies virus vectors as vaccines may be promising for vaccination in animals counteract SARS. Kapadia et al. 85 have created an attenuated vesicular stomatitis virus vector that encodes the SARS-CoV spike. Mice vaccinated with vesicular stomatitis virus S created SARS-CoV-neutralizing antibody and it can control a challenge with SARS-CoV performed at 1 month or 4 months after a single vaccination. Furthermore, by passive antibody transfer experiments, those authors explained that the antibody react induced by the vaccine was enoughfor control of SARS-CoV infection7.
COVID-19 Treatment Methods
There is currently no antiviral or vaccine treatment for COVID-19. Given that there has been tremendous progress in the field of traditional medicine worldwide, we can use traditional medicine like China to treat COVID-19, and on the other hand, we can pay attention to biological control.The production of safe and stable vaccines is a major challenge, and vaccines will ideally be a wide range, and research and development of new drugs is a very long process. In such an epidemic, scientists were unable to produce new drugs in accordance with traditional principles. So, there is another option: a regular and extensive screening of available drugs to find out if they have an effect on the COVID-19. In addition, pitavastatin, perampanel, and praziquantel may be moderately active against COVID-19. According to previous studies, an anti-HIV drug called Kaletra (consisting of two protease inhibitors, ritonavir and lopinavir) has therapeutic effects on SARS. Morse has recently been recommended by the National Health Commission of the People's Republic of China for the treatment of pneumonia of COVID-19. Other medications, including ribavirin, interferon, and corticosteroids used for patients with SARS and Morse, have also been suggested for COVID-19. This effectiveness of these drugs is discussed for COVID-19.Other antiviral drugs, such as FAD-approved drugs such as penciclovir, nitrazine, nalfamusta, chloroquine and two antiviral drugs with a wide range of redexivir and favivir with effects on cytotoxicity, virus function and rate of coronavirus infection. Good effects with invitro have been described in uncoating inhibition and post-translational modification in newly synthesized proteins, particularly glycosylation inhibition in many viruses, including immunodeficiency virus (HIV). Early clinical studies also suggest that chloroquine alone or in combination with antiviral agents may play an important role in the treatment of HIV infection. Recent results have shown that remdesivir and chloroquine are effective in controlling the infection of the COVID-19 in the laboratory and can be evaluated in COVID-19 disease. In addition to one case of COVID-19 pneumonia with a promising clinical response to remdesivir and two clinical trials in China, more clinical trials than remdesivir are needed to confirm its therapeutic efficacy. Remdesivir has also been shown to be a promising antiviral drug against multiple RNA viruses in cultured cells, and in mice and inhumane models (NHP). Remdesivir is currently undergoing clinical trials for the treatment of Ebola virus infection. Remdesivir was used in the treatment of patients with COVID-19 and achieved good results. It is promising that these compounds could cure the COVID-19. In addition, the Fifth Edition of Infectious Diseases Prevention and Control (IPC) states that patients with severe and critical illnesses can be treated with plasma recovery plasma symptoms are therapeutic. The most common complication in patients with the COVID-19 was acute respiratory distress syndrome (ARDS), followed by anemia, acute heart damage, and secondary infections. Therefore, empirical antibiotics, oseltamivir, and systemic corticosteroids are often used for treatment. According to recent reports, more than 85% of patients with antiviral drugs, including oseltamivir (75 mg every 12 hours orally), ganciclovir 0.25 g every 12 hours intravenously, and lopinavir / ritonavir tablets (100-400 mg twice daily) they receive oral administration. Empirical antibiotics are prescribed for 90% of patients in three reports, and according to one study, 15 patients (15%) received antifungal drugs. Five cases (5.1%) of concomitant bacterial infection (1 person) or Candida (4 patients) were reported among 99 patients in one study, and 4 cases (9.8%) of secondary bacterial infections were reported in 41 patients in another study, Although intravenous immunoglobulin and systemic steroids have been used in several reports, their efficacy and side effects are unknown. The new sulfon, in theory, appears to show the potential for antiviral activity against COVID-19. Lu hypothesized that in addition to antiviral and antibiotic interactions, neuraminidase inhibitors, RNA synthesis inhibitors, and traditional Chinese medicine could also be used to treat COVID-19. However, confirmation of the efficacy of these drugs still requires clinical studies. In the absence of effective treatments, the best way to deal with GERD is to control the sources of infection. Strategies include early detection, reporting, isolation and quarantine, and supportive therapies; timely dissemination of epidemic information and maintaining social order is also very effective and useful. For individuals, protective measures, including promoting personal hygiene, wearing medical masks, adequate rest, and proper ventilation of rooms, can effectively prevent the onset of COVID-19.
Caution
Clinical use of glucocorticoids to control coronavirus-2019 pneumonia should be avoided with the aim of regulating cytokine production and inflammatory response and preventing lung damage19.
The use of polymerase virus inhibitors has been used as a successful treatment. A short RNAi was used that targeted RdRp and showed a significant reduction in SARS-CoV plaque formation in Vero-E6cells. However, such an approach could affect the entire 1a / ab protein and not specific to RdRp2. Furthermore, there is clear evidence that recombinant IFN alpha or beta IFN therapy can inhibit SARS-CoV proliferation in laboratory conditions and, most importantly, the severity of SARS-CoV infection within the body. Reduce. Although the antiviral activity of type IFNs in SARS-CoV infections has been clearly demonstrated, it remains to be seen whether significant production of type IFNs can be achieved on coronavirus infection and how this is possible. It is unknown at this time what he will do after leaving the post. It is noteworthy that neither macrophages cDCs fibroblasts nor epithelial lung cell cells which are unable to detect symptomscan type IFN I response to SARS-CoV. Overall, the results of this study show an insightful view of immunopathogenesis of coronavirus-related diseases by showing the isolated role of pFC-derived IFNs for primary viral control. Following this path, for example through dedicated TLR agonists, may open up new avenues for the treatment of COVID-19. In fact, stimulation of TLR3 in the vaginal mucosa can protect mice from the herpes simplex virus challenge through the mucosal route. In a clinical setting, systemic administration of a TLR7 agonist has potent antiviral effects against the hepatitis C virus with a significant reduction in plasma virus5. According to one hypothesis, the first treatment protocols for SARS patients included steroids, which aimed to moderate the intensified cytokine response, as did the treatment of acute non-viral distress syndrome. However, treatment of SARS infection has been ineffective. Treatments were based on the administration of antibacterials (to prevent secondary bacterial infections) and steroids (to regulate cytokine deregulation) along with ribavirin (a broad-spectrum analogue nucleoside with antiviral activity). Currently, there is no antiviral treatment for SARS. Attempts have been made to study the sensitivity to in vitro conditions to different compounds with potential anti-SARS activity. However, many conflicting results have been reported from various laboratories, making it difficult to reach an international agreement on anti-SARS strategies. Use of antiviral antibodies, systemic inhibitors, proteinase inhibitors, calpin inhibitors, virus protease inhibitors type 1 Human safety (inhibition) Proteinase type 1), analog nucleoside (such as ribavirin), interferons, and short-interference RNAs have been reported. Plasma donated from patients who had recovered from SARS was used as immunotherapy for SARS patients. If plasma is in the process of prescribing plasma SARS, it appears to have beneficial effects. These studies suggest that SARS immunoglobulin containing high titers of SARS-CoV neutralizing antibodies could be used for possible future outbreaks. The protective effect of several human SARS-CoV neutralizing antibodies on several human monoclonal bodies has recently been demonstrated using various animal models. It should be noted that although the use of SARS-CoV neutralizing antibodies may be promising, the entry of SARSCoV by antibodies is increased. Interestingly, human antibodies that neutralize quasi-lentivirus viruses that neutralize a type of SARS-CoV generated by most human SARS-CoVs increase the entry of lentivirus independently of glioprotein S palms. The effect of ribavirin on cell culture has been studied by several cell groups by several groups and is still controversial. In general, it appears that depending on the cell type, ribavirin may inhibit SARS-CoV proliferation, but usually at concentrations higher than the average plasma in the treated individuals. There is limited experience with IFN treatment in SARS patients. Treatment with IFN-alphacon-1 resulted in faster clearance. Alpha and beta IFN activity against SARS-CoV in animals has been investigated using sinomolgos masks. Treatment of IFN-alfa-2b Pegylated before SARS-CoV infection protects significant macaques from SARS challenge. Many studies have reported the effects of IFN on SARS-CoV proliferation in laboratory conditions. The antiviral potential of IFN alpha, beta, and gamma has been studied in cell culture, where beta IFN is the strongest inhibitor of SARS-CoV. Following this first report, IFN reported various effects of alpha subgroups and human leukocytes of IFN alpha against SARS reproduction. Gamma IFN gamma has little activity against SARS-CoV in Conditions in vitro. However, IFN beta and gamma may act simultaneously against SARS-CoV infection in laboratory conditions. Short-interference RNAs that prevent the expression of SARSCoV genes have been shown by several groups using different cell categories and also in the Makai model with promising results. Although potential anti-SARS agents are being identified using cell lines as well as SARS animal models, the development of SARS therapies that can be administered to humans quickly and safely in the event of an outbreak based on a better understanding of SARS pathogenesis7. Previous experience has shown that combining ribavirin antiviral drug with interferon alpha-2b can prevent the multiplication of the virus in infected cells in the cell culture medium8.
Health Education Against COVID-19
Coronavirus disease (COVID-19) has spread rapidly in Asia, Europe, the Middle East and the Americas. Considering the recent outbreak of COVID-19, some precautionary measures have been announced, including campus class suspensions. Nursing campus courses have also been suspended, and there may be a learning gap between hand hygiene theory and clinical training for nursing students. A virtual classroom education approach may help address the learning gap by providing ongoing theoretical strengthening of hand hygiene during clinical nursing training86. This editorial offers a 3-step virtual classroom education approach to support nursing educators87 in online theoretical hand hygiene enhancement88. March 2020, it seemed as if my world was turned upside down. I opened an email from my medical school dean of student affairs stating: Match Day, canceled. The day that would determine where we trained as postgraduate residents, the day my class had worked towards for 4 years, canceled due to this new virus that we seemed to know so little about: COVID-19 89. "All hands on deck, we need to move everything online as quickly as possible". This is the message that has been ringing out across universities worldwide over the last couple of months as the Covid-19 pandemic has taken hold. Anatomists have responded to this call enthusiastically and creatively, ensuring that their students have resources available to continue and support their learning as they are forced to study remotely and exercise social distancing and where necessary self-isolation. Despite the seriousness of the health situation, this new drive to move to online learning provides the necessary opportunity to assess our future approach to curriculum design and delivery for the anatomical sciences. This pause to reevaluate is necessary as online learning, offers up refreshing and pedagogically innovative approaches to facilitate learning. For many courses and programs however, it is not the long-term silver bullet solution to all learning provision and has consequences that must be considered90. Preparing for public health emergencies is an ongoing process and involves a variety of approaches and tools. Tabletop exercises are one of the tools designed to simulate the emergence of a public health emergency and address some or all of the phases of emergency management: mitigation, preparedness, response, and recovery. They typically are designed to include participation of stakeholders from diverse and complementary backgrounds, including command, operations, logistics, planning, and finance. Effective tabletop exercises provide a plausible scenario that requires cooperation and communication from these functional areas. Tabletops also require forward thinking and planning in a variety of scenarios. When a public health emergency occurs, decision makers may be overwhelmed with decisions that need their immediate attention91. Dental institutions in the United States are reeling from the consequences of the novel SARS-CoV2 coronavirus, the causative agent of CODIV-19. As oral health care providers, we have been trained on prevention of aerosol transmissible diseases, but we are still grappling with many unknown factors regarding COVID-19. While the Centers for Disease Control and Prevention (CDC), American Dental Association (ADA), and local state agencies are releasing updates on guidelines for dentists and patients, no official information exists for dental institutions on how to effectively follow the recommended guidelines including “shelter in place”with social distancing to protect students, faculty, staff, and patients, and still ensure continuity of dental education92. The COVID-19 pandemic is a huge challenge to education systems so must be offered guidance to teachers, institutional heads, and officials on addressing the crisis. What preparations should institutions make in the short time available and how do they address students' needs by level and field of study? Reassuring students and parents is a vital element of institutional response93.
Conflicts of Interest
None of the authors has any conflict of interest in this manuscript.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
I sincerely thank the Open Access Pub team for encouraging and collaborating with me and revised the manuscript, so I can submit a review article for The Novel Coronavirus 2019 (COVID-19): A narrative review. Also I would like to thank Dr. Morteza Motazakker Associate Professor of Medical Virology Department of Microbiology and Virology, School of Medicine and Patient Safety Research Center of Urmia University of Medical Sciences for his advice.
References
- 1.Ryu Sukhyun, Byung C C. (2020) An interim review of the epidemiological characteristics of 2019 novel coronavirus. , Epidemiol Health 42, 4.
- 2.Benjamin W, Peter Chamberlain Neuman, Bowden Fern, Joseph J. (2014) Atlas of coronavirus replicase structure. Virus Research. 194, 49-66.
- 3.Hui LUO, Qiao-ling TANG, Ya-xi SHANG, Shi-bing LIANG, Ming YANG et al. (2020) Can Chinese Medicine Be Used for Prevention of Coronavirus Disease2019(COVID-19)? A Review of Historical Classics, Research Evidence and Current Prevention Programs. , Chin J Integr Med 1-8.
- 4.Simmonds P, Adams MJ BM, Breitbart M, Brister J R, Carstens E B et al. (2017) Consensus statement: Virus taxonomy in the age of metagenomics. , Nature Reviews Microbiology 15(3), 161-8.
- 5.Cervantes-Barragan Luisa, Zu¨st Roland, Weber Friedemann, Spiegel Martin, Karl S Lang et al. (2007) Control of coronavirus infection through plasmacytoid dendritic-cell–derived type I interferon. , Blood 109(3), 1131-37.
- 6.Cornelia C, Lane TE Bergmann, Stephen A, Stohlman. (2006) Coronavirus infection of the central nervous system: host-virus stand-off. , Nature Reviews Microbiology 4, 121-32.
- 7.Susan R Weiss, Navas-Martin Sonia. (2005) . Coronavirus Pathogenesis and the Emerging Pathogen Severe Acute Respiratory Syndrome Coronavirus MICROBIOLOGY AND MOLECULAR BIOLOGY REVIEWS 69(4), 635-64.
- 8. (2013) . Management of Infectious Diseases 2013 coronavirus Hcov-EMC. In: Ministry of Health TaMED, editor. Tehran-Iran: Minister of Health of the Center for Management of Infectious Diseases 2013 coronavirus Hcov-EMC; .
- 9.Bagheri Sina, Seyed M P, Arash Ghalyanchi Langeroudi. (2019) Detection and Molecular Characterization of Gammacoronavirus in Quail Population in Iran Iranian. , Journal of Veterinary Medicine 13(3), 251-9.
- 10.Sharif Saeed, SitiSuri A, Mohd H, AbdulRahman O, Nazariah A Zeenathul et al. (2010) Diagnostic Methods for Feline Coronavirus: A Review. Veterinary Medicine International. 1-8.
- 11.Valerian V, Eugene V Dolja, Koonin. (2018) Metagenomics reshapes the concepts of RNA virus evolution by revealing extensive horizontal virus transfer. , Virus Res 244, 36-52.
- 12.David M, Arcady R Kristensen, Valerian V Mushegian, Eugene V Dolja, Koonin. (2010) New dimensions of the virus world discovered through metagenomics. , Trends Microbiol 18(1), 11-9.
- 13.Farnoosh Gholamreza, Gholamhossein A, RHZ Seyed, Ruhollah D, Alireza J F. (2020) Understandingthe2019-novel Coronavirus (2019-nCoV) and Coronavirus Disease (COVID-19) Based on Available Evidence -A Narrative Review. , Journal of Military Medicine. January 22(1), 1-11.
- 14.Simmonds P, Adams MJ BM, Breitbart M, Brister J R, Carstens E B et al. (2017) Consensus statement: Virus Taxonomy in the Age of Metagenomics. , Nature Reviews Microbiology 15(3), 161-8.
- 15.Stavrinides J, Guttman D S. (2004) Mosaic evolution of the severe acute respiratory syndrome coronavirus. , J Virol 78, 76-82.
- 16.Goebel S J, Taylor J, Masters P S. (2004) The 3_ cis-acting genomic replication element of the severe acute respiratory syndrome coronavirus can function in the murine coronavirus genome. , J Virol 78, 7846-51.
- 18.Chen Y, Liu Q, DG. (2020) Emerging coronaviruses: genome structure, replication, and pathogenesis. , Journal of medical virology
- 19.Farnoosh Gholamreza, Gholamhossein A, RHZ Seyed, Ruhollah D, Farahani A J. (2020) Understanding the 2019-novel Coronavirus (2019-nCoV) and Coronavirus Disease (COVID-19) Based on Available Evidence -A Narrative Review. , Journal of Military Medicine.January 22(1), 1-11.
- 20.Angelini M M, Akhlaghpour M, Neuman B W, Buchmeier M J. (2013) Severe acuterespiratory syndrome coronavirus nonstructural proteins 3, 4, and 6 inducedouble-membrane vesicles. , MBio 4(4).
- 21.Ziebuhr J, Snijder E J, Gorbalenya A E. (2000) Virus-encoded proteinases and pro-teolytic processing in the Nidovirales. , The Journal of General Virology 81, 853-79.
- 22.Donaldson E F, Graham R L, Sims A C, Denison M R, Baric R S. (2007) Analy-sis of murine hepatitis virus strain A59 temperature-sensitive mutant TS-LA6suggests that nsp10 plays a critical role in polyprotein processing. , Journal of Virology 81(13), 7086-98.
- 23.Imbert I, Guillemot J C, Bourhis J M, Bussetta C, Coutard B et al. (2006) A second, non-canonical RNA-dependentRNA polymerase in SARS coronavirus. , The EMBO Journal 25(20), 4933-42.
- 24.Miknis Z J, Donaldson E F, Umland T C, Rimmer R A, Baric R S et al. (2009) Severe acute respiratory syndrome coronavirus nsp9 dimerization isessential for efficient viral growth. , Journal of Virology 83, 3007-18.
- 25.Bouvet M, Debarnot C, Imbert I, Selisko B, Snijder E J et al. (2010) In vitro reconstitution of SARS-coronavirus mRNA cap methylation. PLoSPathogens 6(4).
- 26.Snijder E J, Bredenbeek P J, Dobbe J C, Thiel V, Ziebuhr J et al. (2003) Unique and conserved features of genome and proteome of SARS-coronavirus, an early split-offfrom the coronavirus group 2 lineage. , Journal of Molecular Biology 331(5), 991-1004.
- 27.Thiel V, Ivanov K A, Putics A, Hertzig T, Schelle B et al. (2003) Mech-anisms and enzymes involved in SARS coronavirus genome expression. , TheJournal of General Virology 84, 2305-15.
- 28.Saikatendu K S, Joseph J S, Subramanian V, Clayton T, Griffith M et al. (2005) Structural basis of severe acute respiratory syndrome coronavirus ADP-ribose-1-phosphate dephosphorylation by a conserved domain of nsP3. , Structure 13(11), 1665-75.
- 29.Lee H J, C K Shieh, Gorbalenya A E, E V Koonin, Monica NLa et al. (1991) The complete sequence (22kilobases) of murine coronavirus gene 1 encoding the putative proteases and RNA polymerase. , Virology 180, 567-82.
- 31.Ziebuhr J, Thiel V, Gorbalenya A E. (2001) The auto catalytic release of a putative RNA virus transcription factor from its polyprotein precursor involves two paralogous papain-like proteases that cleave the same peptide bond. , J Biol Chem 276, 33220-32.
- 32.Smit J J, Rudd B D, L N W. (2006) Plasmacytoid dendritic cells inhibit pulmonary immunopathology and promote clearance of respiratory syncytial virus. , J Exp Med 203, 1153-9.
- 33.TEea Lane. (1998) Dynamic regulation of α- and β-chemokine expression in the central nervous system during mouse hepatitis virus-induced demyelinating disease. , J Immunol 160, 970-8.
- 34.Zhao Y, Zhao Z, Wang Y, Zhou Y, Ma Y et al. (2020) Single-cell RNA expression profiling of ACE2, the putative receptor of Wuhan 2019-nCov.
- 35.Kamitani W, Huang C, Narayanan K, Lokugamage K G, Makino S. (2009) A two-pronged strategy to suppress host protein synthesis by SARS coronavirus Nsp1protein. , Nature Structural & Molecular Biology 16(11), 1134-40.
- 36.Law A H, Lee D C, Cheung BKY, L H C, AS. (2007) Role for nonstructuralprotein 1 of severe acute respiratory syndrome coronavirus in chemokine dys-regulation. , Journal of Virology 81(1), 416-22.
- 37.Brockway S M, Lu X T, Peters T R, Dermody T S, Denison M R. (2004) Intracellularlocalization and protein interactions of the gene 1 protein p28 during mousehepatitis virus replication. , Journal of Virology 78(21), 11551-62.
- 38.Bost A G, Carnahan R H, Lu X T, Denison M R. (2000) Four proteins processedfrom the replicase gene polyprotein of mouse hepatitis virus colocalize in thecell periphery and adjacent to sites of virion assembly. , Journal of Virology 47(7), 3379-87.
- 39.Egloff M P, Malet H, Putics A, Heinonen M, Dutartre H et al. (2006) Structuraland functional basis for ADP-ribose and poly(ADP-ribose) binding by viral macrodomains. , Journal of Virology 80(17), 8493-502.
- 40.Hagemeijer M C, Rottier P J, de Haan CA. (2012) Biogenesis and dynamics of thecoronavirus replicative structures. , Viruses 4(11), 3245-69.
- 41.A von Brunn, Teepe C, Simpson J C, Pepperkok R, Friedel C C et al. (2007) Analysis of intraviral protein-protein interactions of the SARS coronavirus ORFeome. , PLoS ONE 2(5), 459.
- 42.Deming D J, Graham R L, Denison M R, Baric R S. (2007) Processing of open readingframe 1a replicase proteins nsp7 to nsp10 in murine hepatitis virus strain A59replication. , Journal of Virology 81(19), 10280-91.
- 43.Donaldson E F, Graham R L, Sims A C, Denison M R, Baric R S. (2007) Analy-sis of murine hepatitis virus strain A59 temperature-sensitive mutant TS-LA6suggests that nsp10 plays a critical role in polyprotein processing. , Journal ofVirology 81(13), 7086-98.
- 44.A von Brunn, Teepe C, Simpson J C, Pepperkok R, Friedel C C et al. (2007) . Analysis of intraviral protein-protein interactions ofthe SARS coronavirus ORFeome. . PLoS ONE 2(5), 459.
- 45.Zhou H, SP. (2006) Preferential infection of mature dendritic cells by mouse hepatitis virus strain JHM. , J Virol 80, 2506-14.
- 46.Spiegel M, Pichlmair A, Martinez-Sobrido L. (2005) Inhibition of beta interferon induction by severe acute respiratory syndrome coronavirus suggests a two-step model for activation of interferon regulatory factor 3. , J Virol 79, 2079-86.
- 47.Rempel J D, Murray S J, Meisner J, J B M. (2004) Differential regulation of innate and adaptive immune responses in viral encephalitis. , Virology 318, 381-92.
- 48.Jea Zhou. (2005) Expression of matrix metalloproteinases and their tissue inhibitor during viral encephalitis. , J Virol 79, 4764-73.
- 49.Navas-Martin S, S T Hingley, S R Weiss. (2005) Murine coronavirus evolution in vivo: functional compensation of a detrimental amino acid substitution in the receptor binding domain of the spike glycoprotein.J. , Virol 79, 7629-40.
- 50.Grosse B, Siddell S G. (1994) Single amino acid changes in the S2 subunit of the MHV surface glycoprotien confer resistance to neutralization by S1 subunit-specific monoclonal antibody. , Virology 202, 814-24.
- 51.Thackray L B, Holmes K V. (2004) Amino acid substitutions and an insertion in the spike glycoprotein extend the host range of the murine coronavirus MHV-A59. , Virology 324, 510-24.
- 52.Chang Y J, Liu C Y, Chiang B L, Chao Y C, Chen C C. (2004) Induction of IL-8 release in lung cells via activator protein-1 by recombinant baculovirus displaying severe acute respiratory syndrome-coronavirus spike proteins: identification of two functional regions. , J Immunol 173, 7602-14.
- 53.de Haan CA, M de Wit, Kuo L, Montalto-Morrison C, B L Haagmans et al. (2003) The glycosylation status of the murine hepatitis coronavirus M protein affects the interferogenic capacity of the virus in vitro and its ability to replicate in the liver but not the brain. , Virology 312, 395-406.
- 54.Schelle B, Karl N, Ludewig B, Siddell S G, Thiel V. (2005) Selective replication of coronavirus genomes that express nucleocapsid protein. , J Virol 79, 6620-30.
- 55.An S, Chen C J, Yu X, Leibowitz J L, Makino S. (1999) Induction of apoptosis in murine coronavirus-infected cultured cells and demonstration of E protein as an apoptosis inducer. , J Virol 73, 7853-9.
- 56.Sperry S, Kazi L, Graham R, Baric R, Weiss S et al. (2005) Single amino acid substitutions in nonstructural ORF1b-nsp14 and ORF2a 30kDa proteins of the murine coronavirus MHV-A59 are attenuating in mice. , J Virol 79, 3391-400.
- 57.Snijder E J, P J Bredenbeek, J C Dobbe, Thiel V, Ziebuhr J et al. (2003) Unique and conserved features of genome and proteome of SARS-coronavirus,an early split-off from the coronavirus group 2 lineage. , J Mol Biol 331, 991-1004.
- 59.Cheung C Y, Poon L L, Ng I H, Luk W, Sia S F et al. (2005) Cytokine responses in severe acute respiratory syndrome coronavirus-infected macrophages in vitro: possible relevance to pathogenesis. , J Virol 79, 7819-26.
- 60.Reghunathan R, Jayapal M, Hsu L Y, Chng H H, Tai D et al. (2005) Expression profile of immune response genes in patients with severe acute respiratory syndrome. , BMC Immunol 6(2).
- 61.Huang C, Wang Y LX, Ren L, Zhao J, Hu Y. (2020) Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. The Lancet. 395, 497-506.
- 62.Guan W J, Ni Z Y, Hu Y, Liang W H, Ou C Q et al. (2020) Clinical characteristics of 2019 novel coronavirus infection in China.
- 63.Subbarao K, McAuliffe J, Vogel L, Fahle G, Fischer S et al. (2004) Prior infection and passive transfer of neutralizing antibody prevent replication of severe acuterespiratory syndrome coronavirus in the respiratory tract of mice. , J Virol 78, 3572-7.
- 64.Traggiai E, Becker S, Subbarao K, Kolesnikova L, Uematsu Y et al. (2004) An efficient method to make human monoclonal antibodies from memory B cells: potent neutralization of SARS coronavirus. , Nat Med 10, 871-5.
- 65.Meulen ter, ABB J, Brink E N van den, Weverling G J, Martina B E et al. (2004) Human monoclonal antibody as prophylaxis for SARS coronavirus infection in ferrets. , Lancet 363, 2139-41.
- 66.Xiong S, Wang Y F, Zhang M Y, Liu X J, Zhang C H et al. (2004) Immunogenicity of SARS inactivated vaccine in BALB/c mice. , Immunol Lett 95, 139-43.
- 67.Zhou J, Wang W, Zhong Q, Hou W, Yang Z et al. (2005) Immunogenicity, safety, and protective efficacy of an inactivated SARS-associated coronavirus vaccine in rhesus monkeys. , Vaccine 23, 3202-9.
- 68.Zeng F, Chow K Y, Hon C C, Law K M, Yip C W et al. (2004) Characterization of humoral responses in mice immunized with plasmid DNAs encoding SARS-CoV spike gene fragments. , Biochem Biophys Res Commun 315, 1134-9.
- 69.Wang Z, Yuan Z, Matsumoto M, Hengge U R, Chang Y F. (2005) Immune responses with DNA vaccines encoded different gene fragments of severe acute respiratory syndrome coronavirus in BALB/c mice. , Biochem Biophys Res Commun 327, 130-5.
- 70.Zhu M S, Pan Y, Chen H Q, Shen Y, Wang X C et al. (2004) Induction of SARS-nucleoprotein-specific immune response by use of DNA vaccine. , Immunol Lett 92, 237-43.
- 71.Yang Z Y, W P Kong, Huang Y, Roberts A, B R Murphy et al. (2004) A DNA vaccine induces SARS coronavirus neutralization and protective immunity in mice. , Nature 428, 561-4.
- 72.Zeng F, K Y Chow, C, K M Law, C W Yip et al. (2004) Characterization of humoral responses in mice immunized with plasmid DNAs encoding SARS-CoV spike gene fragments. , Biochem Biophys Res Commun 315, 1134-9.
- 73.Zakhartchouk A N, Liu Q, Petric M, L A Babiuk. (2005) Augmentation of immune responses to SARS coronavirus by a combination of DNA and whole killed virus vaccines. , Vaccine 23, 4385-91.
- 74.Gao W, Tamin A, Soloff A, D’Aiuto L, Nwanegbo E et al. (2003) Effects of a SARSassociated coronavirus vaccine in monkeys. , Lancet 362, 1895-6.
- 75.Zakhartchouk A N, Viswanathan S, J B Mahony, Gauldie J, L A Babiuk. (2005) Severe acute respiratory syndrome coronavirus nucleocapsid protein expressed by an adenovirus vector is phosphorylated and immunogenic in mice. , J Gen Virol 86, 211-5.
- 76.Bisht H, Roberts A, Vogel L, Bukreyev A, P L Collins et al. (2004) Severe acute respiratory syndrome coronavirus spike protein expressed by attenuated vaccinia virus protectively immunizes mice. , Proc Natl Acad Sci USA 101, 6641-6.
- 77.Weingartl H M, Copps J, M A Drebot, Marszal P, Smith G et al. (2004) Susceptibility of pigs and chickens to SARS coronavirus. Emerg Infect Dis. 10, 179-84.
- 78.Weingartl H, Czub M, Czub S, Neufeld J, Marszal P et al. (2004) Immunization with modified vaccinia virus Ankara-based recombinant vaccine against severe acute respiratory syndrome is associated with enhanced hepatitis in ferrets. , J Virol 78, 12672-6.
- 79.Martina B E, B L Haagmans, Kuiken T, R A Fouchier, G F Rimmelzwaan et al.SARS virus infection of cats and ferrets. , Nature 425, 915.
- 80.Corapi W V, R J Darteil, J C Audonnet, G E Chappuis. (1995) Localization of antigenic sites of the S glycoprotein of feline infectious peritonitis virus involved in neutralization and antibody-dependent enhancement. , J Virol 69, 2858-62.
- 81.Corapi W V, C W Olsen, F W Scott. (1992) Monoclonal antibody analysis of neutralization and antibody-dependent enhancement of feline infectious peritonitis virus. , J Virol 66, 6695-705.
- 82.Bukreyev A, E W Lamirande, U J Buchholz, L N Vogel, W R Elkins et al. (2004) Mucosal immunisation of African green monkeys (Cercopithecus aethiops) with an attenuated parainfluenza virus expressing the SARS coronavirus spike protein for the prevention of SARS. , Lancet 363, 2122-7.
- 83.Buchholz U J, Bukreyev A, Yang L, E W Lamirande, B R Murphy et al. (2004) Contributions of the structural proteins of severe acute respiratory syndrome coronavirus to protective immunity. Proc Natl Acad Sci USA. 101, 9804-9.
- 84.Faber M, E W Lamirande, Roberts A, A B Rice, Koprowski H et al. (2005) A single immunization with a rhabdovirus- based vector expressing severe acute respiratory syndrome coronavirus (SARS-CoV) S protein results in the production of high levels of SARS-CoV-neutralizing antibodies. , J Gen Virol 86, 1435-40.
- 85.Kapadia S U, J K Rose, Lamirande E, Vogel L, Subbarao K et al. (2005) Long-term protection from SARS coronavirus infection conferred by a single immunization with an attenuated VSV-based vaccine. , Virology 340, 174-82.
- 86.Korthonen A, Vuori A, Lukkari A, Laitinen A, Perälä M et al. (2019) Increasing nursing students' knowledge of evidence-based hand-hygiene: a quasi-experimental study. Nurse Educ Pract. 35, 104-10.
- 87.Ng Y M, Y A Chi-Wing. (2019) 3-step teaching approach for a blended learning of understanding and avoiding unintentional plagiarism. Nurse Educ Pract. 41, 1-3.
- 88.Ng Y M, Or P. (2020) Coronavirus disease (COVID-19) prevention: Virtual classroom education for hand hygiene. Nurse Education in Practice. 45(102782).
- 89.Theoret C, Ming X. (2020) Our Education, Our Concerns: Medical Student Education Impact due to COVID-19. Medical education.
- 90.Evans D, Bay B, Wilson T, Smith C, Lachman N et al. (2020) Going Virtual to Support Anatomy Education: A. STOP GAP in the Midst of the Covid-19 Pandemic. Anatomical sciences education .
- 91.Wendelboe A, Miller A, Patents D, Salinas L, Miller E et al. (2020) Tabletop exercise to prepare institutions of higher education for an outbreak of COVID-19. , Journal of emergency management (Weston, Mass) 18(2), 183-4.
Cited by (1)
- 1.Nugroho Muhammad Fajar, Suswanta Suswanta, 2020, KENDALA PELAKSANAAN DAN UPAYA PENANGANAN WORK FROM HOME /WFH DI BAPPEDA KABUPATEN GUNUNGKIDUL, Jurnal Ilmu Administrasi: Media Pengembangan Ilmu dan Praktek Administrasi, 17(2), 221, 10.31113/jia.v17i2.590