
Influence of Carbon - Nitrogen Supplements and pH on Growth of Sugarcane Stem rot Pathogen Fusarium Solani NVS671
Abstract
Fusarium solani NVS671 identified from infected sugarcane stem of Co 671 as a new pathogen was subjected to various cultural conditions to understand its physiological profile. In continuation with our previous work, cultural analysis was carried out under in vitro condition by supplementing various carbon and nitrogen sources in Czapek-Dox agar (CDA). Under different hydrogen ion concentrations, it was found that the growth of Fusarium solaniNVS671 was less at pH 4 and pH 10 and could reach up to 5 cm after 7 days of incubation on Potato Dextrose Agar (PDA). It is observed that the pH around 7 to 8 was optimum for the growth of F. solani. Ten different nitrogenous (N) sources and nine different carbon sources were tested on CDA medium to know their effect on the mycelial growth rate and characteristics of the fungus. Among the N sources evaluated, ammonium chloride (7.96±0.11 cm) was found to be most efficient for mycelial growth promotion followed by ammonium nitrate (7.7±0.1 cm) and ammonium sulphate (7.3±0.1 cm). The most preferred carbon source recorded to promote best radial mycelial growth was starch (7.96±0.05 cm) and sucrose (7.93±0.05 cm). Capabilities of using different carbon and nitrogen sources and ability to grow at different pH levels may allow species to adapt to specific soil conditions.This study is important to understand the physiology and metabolite preference of F. solani.
Article Information
- Received
- Accepted
- Published
Academic Editor: Samantha Chandranath Karunarathna, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming, China
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2020 Prittesh Patel
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Corresponding author: Corresponding author: Prittesh Patel, Assistant Professor, C.G. Bhakta Institute of Biotechnology, Uka Tarsadia University, Maliba Campus, Bardoli, Surat, Gujarat 394350, India —
Competing Interests
The authors have declared that no competing interests exist.
Funding
No specific funding statement was provided by the authors.
Data Availability
No data-availability statement was provided by the authors.
Citation:
Introduction
Fusarium solani (Teleomorph: Haematonectriahaematococca) is a widely distributed fungal plant pathogen that infects many different genera of plants 1, 2. In nature, Fusarium solani exist as different strains that exhibit variation in their morphological and cultural characters, pathogenicity and virulence. Due to F. solanidistribution across the environments is varied, it is essential to know which isolate is most virulent within a given species. F. solani is pathogenic to at least 111 plant species spanning 87 genera 3. Recently El Haddadi Errifi et al. reported that Fusarium solani had significantly affected preemergence, post emergence mortality rate and growth parameters in Tetraclinis articulate 4. Present investigation was under taken to study the variability in growth requirement and morphological characters of F. Solani.
F. solani and R. solani are the most important saprophytic pathogens, which develop in both cultured and non-cultured soils, causing the symptoms of damping off and root rot diseases to wide range of vegetable and crop plants including tomato 5. It causes wilt and rot diseases on a wide variety of crops including Cucurbita spp.,Pisum sativum and Phaseolus vulgaris 6, 7, 8. Environmental factors such as temperature, and pH have a large influence on fungal growth 9. Study of effect of hydrogen ion concentration on phytopathogen is important to understand their metabolism and probable correlation with soil pH. If the soil pH is favourable for the growth of phytopathogen, the severity of disease can be high. Similarly variation in carbon and nitrogen sources provided may also influence the growth of fungal phytopathogen. Despite the immense agronomic importance of this fungus, change in pH and the pattern of utilization of the substrate have not been worked upon. In present study, we analysed effect of different pH, carbon and nitrogen substrates in PDA and CDA medium on the growth of F.solani NVS671.
Material and Methodology
Fusarium solani NVS671 was identified as a pathogen causing stem rot in sugarcane. In our previous study we have reported that F.solani NVS671 is a new pathogen found to causing stem rot in sugarcane 10. Seven-day-old active culture of F.solani NVS671was used for this study.
Effect of pH (Hydrogen ion Concentration) on Fusarium Solani
Effect of pH on mycelial growth was studied by growing F.solaniNVS671 on PDA mixed with an appropriate volume of the buffer to give desired pH in medium. The pH was obtained over the ranges 4-10 with citric acid or NaOH. The PDB medium and the buffers were autoclaved separately and aseptically mixed during cooling. Then medium was sterilized in autoclave at 120°C for 15 min. A five mm mycelial disc was transferred from the margin of the 7-day-old growing colony, to the PDA plates. After inoculation, Petri dishes were incubated at 28±2°C. The diameter was recorded in Centimetres in two directions at right angles to each other, and then average colony diameter in was calculated and recorded. Measurement of growth was made at 3rd, 5th and 7th day after incubation (DAI). The experimental design was a random complete block (RCB) with three replicate plates of each isolate for each pH treatment.
Effect of Nitrogen Source on Fusarium Solani
To scrutinize the effect of different carbon and inorganic nitrogen sources on growth and cultural characteristics of fungal pathogen Czapek Dox agar (CDA) media were used. All the ingredients with required quantity needed to prepare final amount were weighed and added. Nitrogen sources were added in equal amount to all the individual media. Czapek media without nitrogen source were served as control. Various nitrogen sources such as Potassium Nitrate, Sodium Nitrate, Ammonium Metavanadate, Calcium Nitrate, Cobalt Nitrate, Ammonium Fluoride, Ammonium Chloride, Ammonium Nitrate, Ammonium Oxalate, and Ammonium Sulphate were used at 0.2% concentration.
Effect of Carbon Source on Fusarium Solani
Various carbon sources were weighed and added to the basal medium. A 20 mL of sterilized medium was poured into sterilized Petri plates (90 mm diameter). Inoculations were made with cultural discs (5mm) from seven days old active culture of F. solaniNVS671. Inoculated Petri plates were incubated at room temperature and average colony diameter was recorded after 3, 5 and 7 days. Three replications of each nitrogen and carbon source were maintained.
Statistical Analysis
The experiment was carried out following RCB with three replicates of each treatment. All experimental data of C and N source were subjected to statistical analysis by standard deviation. In case of pH analysis the significance of differences between the treatments was evaluated by one way analysis of variance (ANOVA) at the significance level of 95 %.
Result & Discussion
A total of seven different pH values were evaluated for studying the growth of Fusarium solaniNVS671. Mycelia can grow only within a definite pH range and metabolism is often affected by pH 11. Under acidic condition F. solani growth was very slow and could reach up to 5 cm after 7 days of incubation. It is observed that the optimum pH for the growth of F. solani was 7 to 8 (Figure 1). Further increase in pH had shown reduced growth of phytopathogen. Our data are in accordance with Arunakumara et al., (2017) who reported significant differences in the growth of Fusarium solani at different pH levels 12. Khan et al., (2011) reported that maximum growth of Fusarium oxysporumf. sp. ciceri mycelial mat was observed at pH 7.0 13.
Figure 1. Growth of F. solani NVS671 at different pH. Values are given as Mean growth in Cm ±SEM (n= 3) and considered to be significantly different at P< 0.05.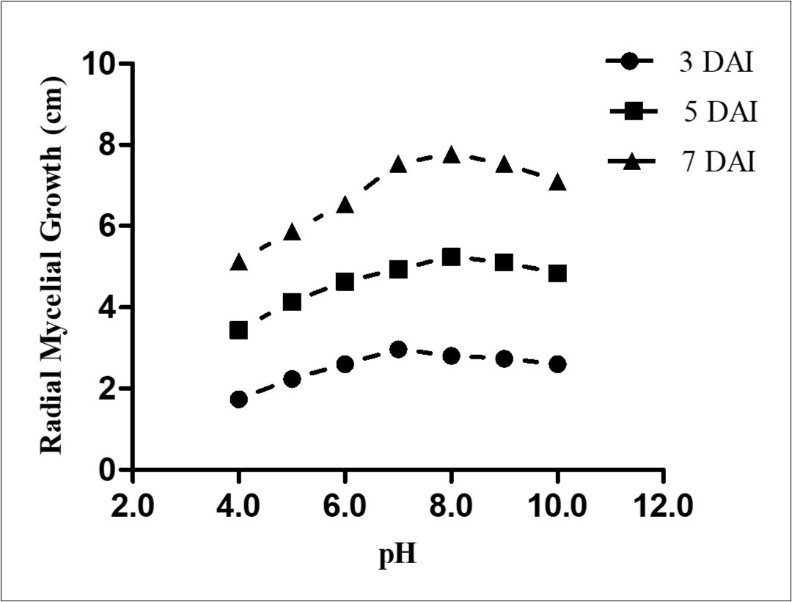
Download figure
It is well known that supplementation of different nitrogen sources in media affect the micro-organism growth. In present study, ten different nitrogenous sources were tested on CDA medium to know their effect on the mycelial growth rate and characteristics of the fungus. All the nitrogen sources had support good mycelial growth at 1.5gm/L concentration except Ammonium Metavanadate and cobalt nitrate. By measuring the radial mycelial growth on CDA, ammonium chloride (7.96±0.11 cm) was found to be most efficient for growth promotion followed by ammonium nitrate (7.7±0.1 cm) and ammonium sulphate (7.3±0.1 cm). Table 1. There was no significant difference among potassium nitrate sodium nitrate and calcium nitrate in efficiency to promote the mycelial growth. Although there was good mycelial growth on ammonium and nitrate, their ability to use different forms of nitrogen are varies. Kamble (2011) reported the best nitrogen source for the growth of Fusarium solani causing rhizome rot of Ginger was Calcium nitrate 14. Ammonium Metavanadate was less efficient as compare to other nitrogen sources and there was negligible growth on CDA media supplemented with it. There was no growth on cobalt nitrate, which indicates that fungus could not able to utilize it. It is possible that cobalt might have toxic effect on F. Solani.
Table 1. Growth of F. solani NVS671 on different nitrogen source (Values represent Mean ± Standard Deviation) (n= 3)| Nitrogen Source | Radial Growth in Centimetre | Mycelial Morphology | ||
|---|---|---|---|---|
| 3 DAI | 5 DAI | 7 DAI | ||
| Potassium Nitrate | 3.0±0.1 | 5.03±0.05 | 6.5±0.1 | White, Uneven, Thin, Sparse growth |
| Sodium Nitrate | 2.93±0.15 | 4.7±0.1 | 6.26±0.11 | Dull White, Uneven, Thin, Sparse growth |
| Ammonium Metavanadate | 0.13±0.05 | 0.23±0.05 | 0.36±0.05 | Dark Grey, Dense growth at centre |
| Calcium Nitrate | 2.26±0.25 | 4.2±0.1 | 5.93±0.15 | White, Uneven, Thin, Scanty growth |
| Cobalt Nitrate | 0 | 0 | 0 | - |
| Ammonium Fluoride | 0.16±0.05 | 0.33±0.05 | 0.5±0.1 | Dark Grey, Dense growth at centre |
| Ammonium Chloride | 3.3±0.2 | 5.6±0.1 | 7.96±0.11 | Dull White, Even, Thin, fluffy growth |
| Ammonium Nitrate | 3.03±0.05 | 5.4±0.1 | 7.7±0.1 | White, Uneven, fluffy growth |
| Ammonium Oxalate | 1.26±0.15 | 2.63±0.15 | 3.7±0.1 | Light Grey, Even, Scanty growth |
| Ammonium Sulphate | 2.96±0.20 | 5.56±0.15 | 7.3±0.1 | White, Uneven, Dense growth at centre |
Carbon is one of the most important nutrients required by living organisms for its growth. In present study, total nine different carbon sources were tested on CDA medium to observe their effect on the mycelial growth rate and characteristics of the fungus. All the nine kinds of carbon sources tested were found suitable for fungus growth. The efficiency of individual carbon sources could be divided into three categories. The first category which recorded to promote best radial mycelial growth included was starch (7.96±0.05 cm) and sucrose (7.93±0.05 cm). The capability of the fungus to utilized sucrose and starch was due to the possession of relevant enzymes to hydrolyze them into their component sugars 15. The second category included maltose lactose dextrose and mannitol which showed to efficiently supported growth between 6.9 cm and 7.7 cm on the 7th day. Cellulose was in the third category which was the least satisfactory. The growth of the pathogen in different carbon sources exhibited significant differences among the treatments. Several researchers reported Dextrose as best carbon source for growth of F. oxysporum16, 17. (Table 2)
Table 2. Growth of F. solani NVS671 on different Carbon source (Values represent Mean ± Standard Deviation) (n= 3)| Carbon Source | Radial Growth In Centimetre | Mycelial Morphology | ||
| 3 DAI | 5 DAI | 7 DAI | ||
| Mannitol | 3.13±0.05 | 5.76±0.20 | 7.23±0.11 | White, Uneven, Scanty growth |
| Starch | 3.3±0.1 | 6.33±0.05 | 7.96±0.05 | Dull white, Uneven, Thick fluffy growth |
| Cellulose | 2.13±0.05 | 3.03±0.15 | 4.83±0.05 | Light white, Even, Dense mycelia at centre |
| Sucrose | 3.86±0.05 | 6.53±0.35 | 7.93±0.05 | Dull white, Even, Thin growth |
| Maltose | 3.73±0.05 | 6.83±0.05 | 7.7±0.2 | Light grey, Uneven, Sparse growth |
| Dextrose | 3.8±0.1 | 6.33±0.11 | 7.5±0.2 | White, Even, Scanty growth |
| Fructose | 3.03±0.05 | 5.13±0.05 | 6.96±0.11 | White, Uneven, Thick fluffy at centre |
| Lactose | 3.46±0.05 | 6.26±0.25 | 7.7±0.1 | Dull white, Even, Fluffy growth |
| Xylose | 3.03±0.05 | 5.53±0.05 | 6.9±0.2 | White, Uneven, Thin growth |
Conclusion
This study is essential to recognize the physiology and metabolite preference of Fusarium solani. Capabilities of using different N and C sources and ability to grow at different pH levels may allow species to adapt to specific soil conditions. Future research should be directed toward the study of the enzymatic activity of Fusarium solani. In this way, it will be possible to obtain a more complete picture of the characteristics of Fusarium solani.
References
- 1.Bogale M. (2009) DiverseFusariumsolaniisolates colonize agricultural environments in Ethiopia. European journal of plant pathology. 124(3), 369-378.
- 2.A Y Rossman. (2013) Genera in Bionectriaceae, Hypocreaceae, and Nectriaceae (Hypocreales) proposed for acceptance or rejection. , IMA fungus 4(1), 41-51.
- 3.Singh R, Singh U, Kohmoto K. (1995) Pathogenesis and host specificity in plant diseases. Vol. III. Viruses &viroids, Elsevier Science , Oxford, UK .
- 4.R El Haddadi. (2019) First report of Fusariumsolani causing damping-off disease on Tetraclinisarticulata seedlings. , Plant Cell Biotechnology and Molecular Biology 20(23), 1106-1114.
- 5.Amira M. (2011) Assessment of antifungal activity ofRumexvesicariusL. andZiziphusspina-christi(L.) Willd Extracts against two phytopathogenic fungi. , African Journal of Microbiology Research 5, 1001-11.
- 6.Hawthorne B, Rees-George J, Broadhurst P. (1992) Mating behaviour and pathogenicity of New Zealand isolates ofNectriahaematococca(Fusariumsolani). , New Zealand Journal of Crop and Horticultural Science 20(1), 51-57.
- 7.H D VanEtten. (1978) Identification of additional habitats of Nectriahaematococca mating population VI. , Phytopathology 68, 1552-1556.
- 8.Li D. (1995) TheFusariumsolanigene encoding kievitone hydratase, a secreted enzyme that catalyzes detoxification of a bean phytoalexin. Molecular Plant Microbe Interactions. 8(3), 388-397.
- 9.R S Yadav. (2014) Effect of different cultural condition on the growth ofFusariummoniliformecausing bakanae disease. , European Journal of Molecular Biotechnology 2, 95-100.
- 10.Patel P.. (2019)FusariumsolaniA New Pathogen that Causes Stem Rot of Sugarcane in South Gujarat, India. National Academy Science Letters: 1-4.
- 11.S W Kim. (2003) Optimization of submerged culture process for the production of mycelial biomass and exo‐polysaccharides byCordycepsmilitarisC738. , Journal of applied microbiology 94(1), 120-126.
- 12.Arunakumara K, Satyanarayana C. (2017) Impact of abiotic and nutritional factors on growth ofFusariumsolanicausing root rot of okra. , The Journal of Rural and Agricultural Research 17(2), 74-77.
- 13.Khan I. (2011) Effect of different media and environmental conditions on the growth ofFusariumoxysporumf. sp. ciceri causingFusariumwilt of chickpea. , Int. J. Sci. Nature 2(2), 402-404.
- 14.Ramteke P, Kamble S. (2011) Physiological studies inFusariumsolanicausing rhizome rot of ginger (ZingiberofficinaleRosc.). , The Bioscan 6(2), 195-197.
- 15.V K Ravat, Basu A. (2019) Supplement of carbon and nitrogen nutrition towards assessment of biomass and virulence of rice fungal pathogen and endophytes. , Journal of Pharmacognosy and Phytochemistry 8(2), 285-295.
Cited by (6)
This article has been cited by 6 scholarly works according to:
Citing Articles:
H. Akram, Shoaib Hussain, Talib E. Butt - Trends in Horticulture (2025) Semantic Scholar
Karantin i zahist roslin (2024) OpenAlex Crossref
Environmental Pollution (2022) OpenAlex Crossref
H. Akbarian, F. Jalali, Mohammad Gheibi, Mostafa Hajiaghaei-Keshteli, Mehran Akrami et al. - Environmental Pollution (2022) Semantic Scholar
Asian Journal of Microbiology Biotechnology and Environmental Sciences (2022) OpenAlex Semantic Scholar
Revista de Investigaciones Altoandinas - Journal of High Andean Research (2021) OpenAlex Semantic Scholar