Use of Immune Modulator Interferon-Gamma to Support Combating COVID-19 Pandemic
Abstract
While the COVID-19 pandemic has raised concerns about the future of people worldwide, it has made it necessary to take measures with high economic costs, including quarantine. We consider it is more logical for some scientists to investigate time-saving treatment options until vaccination studies, which are started to be studied rapidly, are accomplished or specific antiviral agents are found. In this context, treatment combinations of one or more of the immune modulators known as cytokines, which can stimulate or accelerate the immune system, should be tried. In our opinion, although such options are not as effective as specific treatments such as vaccines, such options will offer highly effective alternatives in times of emergency. For this reason, we found it appropriate to make a reminder by preparing a broad review about interferon gamma, which is an antivirus and is an immunomodulator and which plays a critical role in humoral and cellular immunity.
Article Information
- Received
- Accepted
- Published
Academic Editor: Sasho Stoleski, Institute of Occupational Health of R. Macedonia, WHO CC and Ga2len CC, Macedonia.
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2020 Fatih Ozcelik, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Corresponding author: Fatih Ozcelik, University of Health Sciences Turkey, Sultan, 2, Abdulhamid Han Training Hospital, Department of Medical Biochemistry, 34668, Istanbul, Turkey —
Competing Interests
The authors have declared that no competing interests exist.
Funding
No specific funding statement was provided by the authors.
Data Availability
No data-availability statement was provided by the authors.
Citation:
Introduction
Today, there has been a significant increase in the geriatric human population due to prolonged lifespan. The number of people who have weakened naturally and acquired (adaptive) immune system, which creates antiviral defenses, has also increased. This creates a favorable environment for opportunistic microorganisms such as bacteria, fungi and viruses, in which they can live and multiply. In addition, super-infections caused by many microorganisms due to developing serious resistance against existing antibiotics enrich this environment 1, 2, 3. As if these were not enough, the fact that many microorganisms, which normally do not pose a significant threat to humans, begin to cause disease by mutating, further increases the extent of the danger. Due to climate change and intervention in habitats, microorganisms with a suitable genotype for mutation and adaptation to the changing environment have led to the emergence of fatal diseases of which specific treatment is not yet known and leaves the available treatment options inadequate 4, 5. The novel coronavirus 2019 (2019-nCoV or COVID-19) pandemic, one of the biggest epidemics of the last century, is the result of such a process. Deaths related to this pandemic cause great concern worldwide and cause great economic losses due to quarantine measures taken. The COVID-19 pandemic causes immediate decisions with insufficient risk assessment and limited information 5. In this period, when no successful results have been obtained from the specific drug and vaccine studies that have been rapidly initiated, it will be very useful to investigate options that stimulate and strengthen the immune system against COVID-19 virus. In this context, we think that the treatment protocols formed by one or more suitable immunomodulating agents, which can stimulate or accelerate the immune system, especially in the elderly population, should be tried. Therefore, we find it useful to review information about interferon-gamma (IFN-γ), known as antivirus and plays a critical role in humoral and cellular immunity with its immunomodulator properties.
Immunomodulators and Impact Mechanisms
The infection only causes disease when the interaction between the host and the microorganism causes damage leading to disrupt of homeostasis. Treatment to prevent infections in the process until today is mostly based on antimicrobial and antiviral drugs. This approach has been particularly successful against bacterial infections. However, the resistance developed by microorganisms has brought immunomodulator therapies to the agenda, bringing the host to the fore. In immunomodulatory therapy, the goal is not to combat the pathogen, but to modulate the host's immune system to provide a quick and adequate response. In other words, the aim is to improve the interaction between the host and the microorganism in favor of the host (Figure 1). Substances that function in this way are called immunomodulating agents. What is expected of immune modulating agents is to induce the immune system delayed in response or to strengthen the low level response 6, 7, 8. Thus, immunomodulators strengthen humoral immunity by stimulating defense against extracellular pathogens (such as viruses and bacteria) and toxins (neutralization, opsonization and complement activation) through antibodies and strengthen cellular immunity by stimulating the T helper (CD4) and T cytotoxic (CD8) response to intracellular pathogens. Immunomodulators that stimulate natural immunity also play a critical role by preventing a non-specific response to the pathogen. They can also provide a broad-spectrum emergency treatment option in the event of a new pathogen-induced epidemic or biological attack such as COVID-19.
Figure 1. The balance between the host and the microorganism. When balance develops against the host, microbial diseases occur. Immune modulating agents can ensure that balance is restored.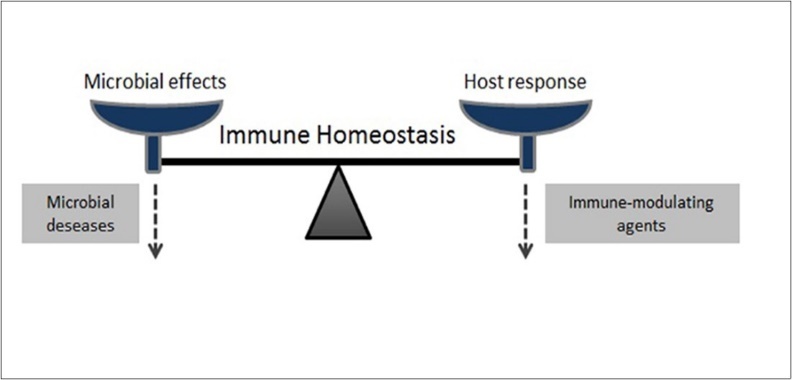
Download figure
In general, immune modulators have less potential to develop treatment resistance compared to treatments targeting specific pathogens. In addition, specific treatments are effective against one or more pathogens or diseases, while immunomodulators can be used as treatment options in the wider disease group. Because their mechanism of action includes multiple effector cells and mediators. For example, Natural Killer (NK) cells activated by immunomodulating agents, by secreting INF-γ, both stimulate the phagocytic activities of macrophages and destroy virus-infected or modified cells and cancer cells. NK cells have inhibitory and activating receptors that tightly regulate their effects to detect abnormal cells and be tolerant to healthy cells. The effectiveness of NK cells depends on the docking of these activating or inhibiting receptors. Docking with NKG2D and CD16 receptors activates NK cells. By activation, NK cells express inhibitory receptors (iNKRs) that specifically bind MHC-I molecules on the surface of healthy cells. In some viral infections and cancer, the expression of MHC-I molecules on the cell surface decreases. Therefore, inhibitory signals of iNKRs receptors are lost. Thus, infected cells become targets for NK cell activation. In relation to this mechanism, many viruses, including some types of EBV, CMV, HIV, and Coronaviruses, have been found to reduce MHC-I expression to avoid T cell responses 6, 9, 10, 11, 12, 13, 14. In studies, Synthetic peptides of MHC proteins (HLA in humans) have been found to show immunomodulatory effects in animals and humans in vitro and in vivo.
Immunomodulators as an Option in Treatment
There is a wide variety of viruses in the world that can be expressed in millions. This wide variety of viruses can cause disease of varying severity in plants, animals and humans. Especially the survival of human and animal hosts depends on the cellular mechanisms of the innate antiviral immune response and the elimination of the virus by virus-specific lymphocytes 15, 16. Therefore, the focus is on the development of broad-spectrum antiviral approaches that can stimulate these mechanisms. Because, such approaches as in the case of IFNs, which activates protective mechanisms for both virus control and elimination, such approaches have the potential to resist different viruses with different disease pathogenesis. They are also quite suitable for prophylaxis and early treatment.
In a recent study, the identification of the crystal structure of the HCoV-EMC protease revealed the idea that a CoV protease inhibitor can block the catalytic region 17. However, there is currently no proven antiviral therapy for HCoV-EMC. Viruses often capture cellular processes that are initiated in response to infection and use the host to replicate. Targeting the cellular response in treatment can inhibit viral replication 18, 19. Also, as with influenza virus infections, immunomodulatory drugs, which reduce the host's excessive inflammatory response, defined as cytokine storm, have the therapeutic benefit. Targeted cytokines have many functions. These functions can be summarized according to the basic molecular groups. One of these groups, interferons exhibit regulation of natural immunity, activation of antiviral properties and antiproliferative effects. Interleukins, another group of cytokines, attract attention due to their growth and differentiation of leukocytes and mostly their proinflammatory effects. Chemokines are chemoattractant for leukocytes, as well as most of them have proinflammatory properties and are involved in the removal of leukocytes from blood to tissue. Colony-stimulating factors stimulate hematopoietic progenitor cell proliferation and differentiation. Tumor necrosis factor (TNF) activates cytotoxic T lymphocytes as well as proinflammatory properties. Taking into account these features, the various anti-inflammatory drugs which are used are approaches for targeting the host response and cytokines, which are in several acute severe infections. Approaches to provide immune modulation include corticosteroids, aspirin, specially selected T lymphocyte clones (e.g., T lymphocyte clones reacted as a result of incubation with various microorganisms), recombinant INF-γ, polyethylene glycol (PEG) -IFN-λ1, some specific antigen receptors (e.g., chimeric specific antigen receptors that have been tested in HIV infection), antigen-specific monoclonal antibodies and some adjuvants, anti-cytokine and anti-chemokine agents, plasma exchange and statins 20, 21, 22, 23. However, the impact of none of these options has been clearly proven. Some even worsened the situation.
Immunomodulators have benefits as well as some negative aspects. The major negatives of immunomodulators are that the response is very complex, its mechanism is not fully understood, it causes undesirable effects in other metabolic processes, it can generate an inflammatory response that is more than expected and cannot be silenced, and can trigger autoantibody production, which can damage itself. These congenital immunomodulators should not be expected to treat the disease alone. They should be considered as special helpers in combination with traditional antiviral and antibacterial treatments. Also, the timing of their use can be very critical. Because, while they show a protective function when used before infection, they can increase the damage in the host when used after infection 6, 7, 24, 25. That is, it is possible that immunomodulators have both positive and negative effects at different stages of the same disease.
Cytokines effective in the natural immune system are TNF, IL1, IL12 and INF. IL2, IL4, IL5, IL17 and INF-gamma are among those that are effective in acquired immunity. This review will focus more on INF-gamma and recombinant INF-γ.
Overview of Interferons as an Immunomodulating Agent
IFNs are glycoprotein-based immunomodulators produced by the immune system as a defense against viruses and included in the class of cytokines. They have autocrine and paracrine activities. There are three types of interferons (alpha (α), beta (β) and gamma (γ)). These interferons are divided into two groups. Interferon α and β forms (type I), are the most abundant and best studied, and can be produced by all cells that are stimulated by the virus. The main function of interferon α is to trigger viral resistance. Interferon-γ is secreted by NK cells and T lymphocytes (Figure 2).
Figure 2. General functions of IFN-α and IFN-γ. IFN-α stimulates viral resistance, whle IFN-γ enhances antigen presentation to strengthen the immune response.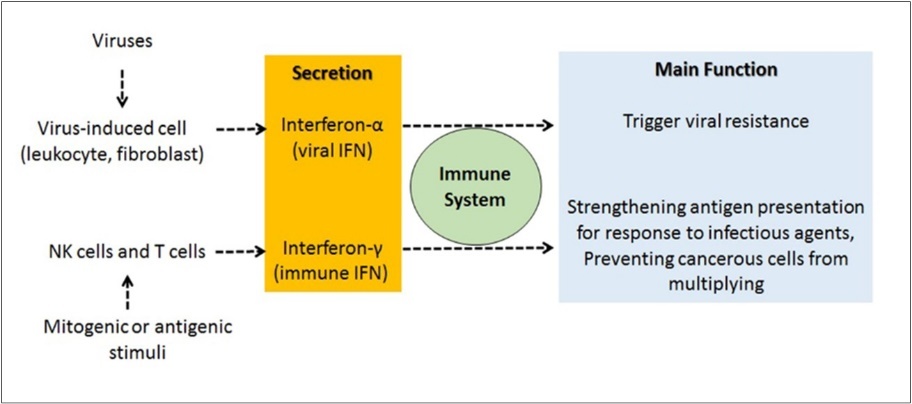
Download figure
In 2003, IFN-lambda (IFN-λ), consisting of 3 cytokines with heterodimeric receptors, was found. It has been determined that it is encoded with 3 different genes called IFN-λ1, -λ2 and -λ3. IFNλs, which consists of these three cytokines, are called type III IFN as a group. Type I IFNs and type III IFNs use the same intracellular signaling pathway, although they are found to produce signals through different receptor complexes 26, 27, 28, 29. Therefore, they show the same biological activities by affecting the same target cells, and both are very similar to antiviral activity. However, the receptors of type I IFNs are expressed in many cells, including leukocytes, while the receptors of type III are largely expressed in epithelial cells. It is thought that Type III IFNs may be useful for use in relation to COVID-19, which is associated with severe acute respiratory syndrome caused by SARS-CoV-2. However, uncertainties remain regarding the exact effects and patient selection 16.
The IFN response represents an early host defense that takes place before the immune response begins. Its main function is to signal the immune system by responding to infectious agents such as bacteria and viruses by strengthening antigen presentation or to prevent the proliferation of cancerous cells 27, 30, 31. There are some proteins that are responsible for the antiviral effects of IFNs. These include RNA-dependent protein kinase (PKR), 2',5'-oligoadenylate synthetase (OAS), RNase L, Mx protein GTPases and RNA-specific adenosine deaminase (ADAR). Their production is stimulated by IFNs. IFNs also stimulate a serial of inducible nitric oxide synthase (iNOS 2) and major histocompatibility complex I and II proteins. All these proteins play an important role in immune response to infections. For this reason, IFNs have been transferred to clinical applications, confirming their use for therapeutic purposes. Many animal viruses found in nature are inducers of IFN. They are also sensitive to the antiviral effects of IFNs 27. Some animal viruses can encode products that antagonize the antiviral response of IFN.
Interferon Type I (Alpha and Beta)
IFNs type 1 play an important role in antiviral immunity. Due to their immunomodulatory properties, IFNs type I is used in the treatment of many diseases. For example, they are used in the treatment of various leukemias, multiple myeloma, Non-Hodgkin lymphoma, Acquired Immune Deficiency Syndrome (AIDS) -related Kaposi Sarcoma, renal cell carcinoma, multiple sclerosis and metastatic carcinoid tumor. In addition, subcutaneous IFN-α and β injections have been used for a long time to treat patients that infected by hepatotropic viruses such as hepatitis B and C 32, 33, 34, 35, 36, 37. Their use in MERSCoV infection has recently been investigated. It has been disclosed that IFN-β1 may be a safe treatment option against COVID-19 in the early stages of infection. The same treatment has been reported to show a different efficiency against MERS-CoV and SARS-CoV viruses 38. In addition, it was emphasized that the findings should be confirmed with larger studies. Contrary to these judgments, the determination that COVID-19 pathology, which mainly produces pulmonary lesions, has similar features to IFN-induced pathologies, increased negative thoughts against this treatment option. In support of this negativity, it has been reported that SARS-CoV-2 can cause tissue damage by over-inducing the IFN-I-mediated antiviral response. Due to clinical data showing that interferons and other inflammatory biomarkers are associated with increased mortality, the hypothesis has been proposed that IFN type I treatment should be limited only to the early stages of infection 39, 40. In late stages, the use of anti-IFN drug has been proposed to alleviate severe course of the pathology consisting of inflammation 41.
As it turns out, each type of interferon has different activities in the immune system and one's deficiency cannot be repaired by the others. For example, mice, which have robust other IFN gene expressions but have defects in the expression of a single IFN-β gene, have been found to be highly susceptible to viral infection 42. This is evidence that IFN-α subspecies or IFN-γ cannot compensate for loss of IFN- γ and IFN- β is required for an effective antiviral response. Type 1 interferons have a wide range of antiviral activity in vitro and are tried in a clinical trial as an option in the treatment of MERS-CoV 38. In many emerging viral infections, IFN type I due to their nonspecific antiviral effects is generally evaluated in combination with other drugs before specific treatments are developed 43, 44. IFN type I, which is secreted by various cell types (especially plasmacytoid dendritic cells) upon the recognition of viral components by pattern recognition receptors (PRR), is among the first cytokines produced during a viral infection. It is recognized by the IFN-α receptor (IFN-αR) found in the cell membrane in most cell types. The fixation of IFN-α on IFN-αR performs phosphorylation of transcriptional factors such as STAT1. They are then relocalized to the nucleus. There, they activate IFN-induced genes (ISG). Most ISGs play a role in inflammation signaling and immunomodulation. They inhibit viral replication and slow cell metabolism. They also stimulate the secretion of cytokines that support the activation of adaptive immunity. ISGs contain PRRs and proteins that make the cell even more sensitive to pathogens 45, 46, 47. These include antivirals that reduce membrane fluidity, prevent viral outflow or membrane fusion and specifically inhibit one step of the viral cycle.
Interferon Type II (Gamma)
Interferons, which can be produced quickly with the warning of viruses or foreign substances, constitute a very important part of body defense. Interferons prevent the reproduction of the virus indirectly, not directly. They stimulate the production of certain proteins, causing the destruction of infected and adjacent cells. Therefore, they prevent the reproduction of the virus. They are also effective in preventing bacterial and parasitic infections. Interferons increase T-lymphocyte activity and cellular breakdown capacity of NK cells while inhibiting B-lymphocyte activation. Recent studies have found that IFNs play an important role in humoral immunity by promoting antibody production of B lymphocytes, and in the stimulation and regulation of CD4 + T cells independent of the antibody. In recent studies, it has been found that IFNs play an important role in humoral immunity by promoting antibody production of B lymphocytes, and in the stimulation and regulation of CD4 + T cells independent of the antibody. It was also found that they contribute to the polarization of T helper 1 (Th1) and T helper 2 (Th2) and mediate antibody-independent functions by secreting different cytokines 48, 49, 50, 51. These INFs have been reported to suppress B10 cells producing regulatory IL-10 and CD4 + T cell mediated hypersensitivity reaction and prevent the induction of autoimmune diseases. It has been reported that they can induce the production of CD4 + and CD8 + regulatory T cells, which control the immune response occurring, as in B40-activated B cells. In addition, a sub-population of innate B lymphocytes, which produces a high amount of IFN-γ, which can promote macrophage activation and facilitate its natural immune response against intracellular bacterial infections, was found in mice exposed to Listeria monocytogenes (LM), Escherichia coli and vesicular stomatitis virus 52, 53, 54, 55, 56. Regardless of antibody production, this innate immunity 57 can be regarded as the memory of the organism that is transmitted by generations against infections. It is worth exploring whether this innate B lymphocyte-mediated immunity can also prevent COVID-19 infection, which can mutate in animals and cause disease in humans. Presumably this congenital B lymphocyte-mediated immunity is ineffective against newly derived infections that the organism has not previously encountered. Presumably this congenital B lymphocyte-mediated immunity is ineffective against newly derived infections that the organism has not previously encountered.
Monocytes, which are cells of the innate immune system, are located on the front line of defense in the body. When they encounter microorganisms that invade the body, they quickly become active. Monocytes can also differentiate into dendritic cells (DCs) known as accessory cells in humans and animals. DCs are cells that offer antigens to the immune system. Their main function is to process the antigen and present it to T cells. Dendritic cells are the precursors of cells that produce natural IFN-α. They are messengers that act between natural and adaptive immune systems. DCs regulate the functions of different types of lymphocytes in response to against pathogens, providing the initiation and control of the adaptive immune response in the T cell. Also, different types of DCs can affect the development, proliferation and activation of B cells. Activated DCs provide differentiation of B cells through cytokines. They can transform antigen-specific B cells into plasma blasts that secrete IgM 58, 59, 60, 61. Studies show that DCs (not NK or T cells) activated by the invasion of pathogens can induce differentiation of B cells.
Interferons (IFNs) have multiple functions in innate and adaptive immune response to eliminate invading viruses. IFN-γ production mechanisms remain uncertain in the innate immune response. IFN- γ, which is thought to be produced by NK cells and CD4 + T cells, may increase innate immunity 62. Considered to be produced mainly by NK cells and CD4 T cells, IFN-γ can enhance innate immunity by stimulating antimicrobial factors or destruction pathways in other immune cells such as macrophages. It causes this by directly inhibiting viral replication and activating the immune system to eliminate viruses. Combined TLR (Toll-like receptor) ligands and stimulation of IL-12 / IL-18 or PMA / ionomycin can trigger B cells to make many cytokines, including IFN-γ in vitro 63, 64, 65, 66. Increased IFN-γ dependent innate immune response is of great importance in host defense against invading pathogens. However, the role of IFN-γ producing B cells in this process is not exactly known. NK cells, which are rapidly activated approximately 14-20 hours after infection, are the main source of innate responses in the first stage of IFN-γ. In this way, activated macrophages maintain the serum IFN-γ level, keeping microorganism growth under control 67, 68, 69. Studies report that innate B cell numbers producing IFN-γ may peak only on day 3 after infection. In summary, innate B cells are not induced very quickly. This suggests that B cells may participate in innate defense against LM after the initial activation of NK cells. This is another important source of IFN- γ for NK cells to complete their functions.
IFN-γ with antiviral activity is a homodimeric glycoprotein with pleiotropic immunological functions. It is an important cytokine that plays a role in stimulating and modulating a range of immune responses 48, 70. IFN-γ is generally secreted by activated T cells and natural killer (NK) cells. IFN-γ can affect many mechanisms in the immune system. Namely, by strengthening antigen presentation, it shows antiviral and antibacterial activity, stimulates macrophage activation and triggers the innate natural immune system. It is also reported that it regulates lymphocyte-endothelial interaction and Th1 / Th2 balance (two types of effector helper T cells caused by differentiation of CD4 + T lymphocytes) and can control cellular proliferation and apoptosis. For these effects to occur, IFN-γ must bind to heterodimeric cell surface receptors (IFN-γR1/2). A complex set of signals that result from binding triggers the cascade, creating the cellular response that leads to regulation of gene expression. Basically, the signaling triggered by binding of IFN-γ is predicted to begin with JAK2 autophosphorylation followed by phosphorylation of JAK1 (Figure 3). The activated JAK1 is thought to phosphorylate IFN-γR1 by providing a site of insertion for STAT1 48, 71, 72, 73, 74, 75, 76. After binding to the receptor site, STAT 1 is believed to be activated by JAK2 through phosphorylation.
Figure 3. IFN-γ signal path. Binding of IFN-γ to IFN-γR1/2 receptors leads to activation of pairs of JAK and STAT transcription factors that overlap with tyrosine phosphorylation. Phosphorylated STAT1 proteins are homodimerized through mutual phosphotyrosine interactions and GAF is formed. Then, the GAF (gamma-activated factor) passes to the nucleus by translocation and connects to a nine-nucleotide consensus sequence known as GAS (gamma-activated site). This region contains more than 200 protein-coding genes.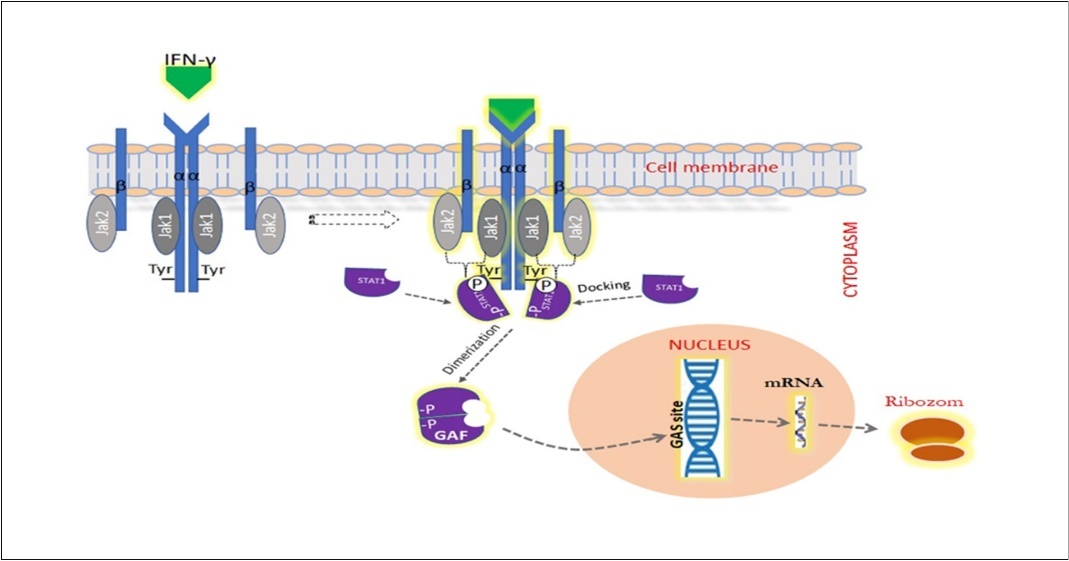
Download figure
IFN-γR1/2 receptor to which IFN-γ binds, consists of α and β (R1 (CD 119) and R2) chains 71, 77. The affinity of IFN-γR1 to IFN-γ is high. However, it cannot produce a biological response to cytokine stimulation alone. Because, there is also need for the β chain, which is important in receptor stability. Some amino acid residues in the cytoplasmic portions of IFN-γR1/2 play a critical role in the activation of intracellular signaling pathways that will deliver the IFN-γ signal from the cell surface to the nucleus. A mutation that will affect these residual amino acids in the IFN--γR1/2 glycoprotein will cause serious defects in the immune system 73, 74, 77, 78, 79, 80, 81, 82. IFN-γR has no intrinsic kinase activity in α and β chains, such as members of the class II family of cytokine receptors (tissue factor, IL-10 ligand binding component, and IFN-α receptor). Therefore, signaling takes place through JAK1 and JAK2, members of the family of protein tyrosine kinases that are structurally related to specific membrane-proximal residues in the cytoplasmic areas of IFN-γR. JAK 1 connects to a region on IFN-γR1 and JAK2 connects to a region on IFN-γR2 73,83, 84, 85, 86. The aggregation of IFN-γR1/2 receptor components with the stimulation of IFN-γ enables the inactive JAKs associated with the cytoplasmic ends of α and β chains to converge towards each other. Clustered JAKs are mutually activated through a series of auto- and transphosphorylation reactions 84, 87. Activated JAKs phosphorylate a specific tyrosine residue near the C-terminal at both ends of IFN-γR1. This pair of phosphorylated tyrosine residue is found in a recognition sequence associated with the domain of STAT1 (member of the latent cytoplasmic proteins signal transducers and activators of the transcription family) defined as SH2. After the STAT1 molecules are placed in their target sequences on the IFN-γR complex, they are phosphorylated from the tyrosine residue by the receptor-associated JAKs. The two phosphorylated STAT1 proteins transform into a homodimeric form through SH2-phosphotyrosine interactions, forming a protein complex called gamma activated factor (GAF). This homodimer is then transported to the nucleus where it binds to a 9 nucleotide consensus sequence known as the gamma activated region (GAS) element 86,88, 89, 90, 91, 92, 93. This binding site has also been detected in the regulatory regions of more than 200 genes. Thus, the signal initiated by IFN-γ can modulate the expression of a large gene sequence to mediate biological functions. Initially, Suppressors of Cytokine Signaling expression is induced by IFN-γ 71, 94. After activating the cells with the IFN-γ signal, their sensitivity to this cytokine decreases and a negative feedback occurs.
Immunological memory based on T helper cell function may be affected by the absence of a functional IFN-γ system. In a study conducted for this purpose, it was found that secondary T cell response could not be obtained when exposed to antigen for the second time in the absence of INF-γR1 receptors (Tau). Like CD4 + T helper cells, CD 8+ cytotoxic T lymphocytes are activated by an antigen presented in the context of an MHC 1 class molecules. MHC class I molecules involved here are expressed everywhere different from MHC class II. Moreover, antigen presentation to the MHC class II pathway can only be induced by IFN-γ, while antigen presentation to the MHC class I pathway can be induced by IFN-γ or IFN-α. Tc1 cells, a subset of dominant CD8 + T lymphocytes, secrete significant amounts of IFN-γ mediating their effector functions 95, 96, 97, 98, 99. In some studies, it has been found that while there is resistance against some viruses (such as herpes virus, lymphocytic choriomeningitis virus and mouse mammary tumor virus) in INF-γ deficiency, it is still susceptible to some viruses (vaccine virus, murine cytomegalovirus and coronavirus). This situation indicates that type I IFNs produced by any cell are necessary for protection against viral infection, but IFN-γ produced only on stimulation of subgroups of specific immune cells with antigen and mitogen is pathogen dependent 62,100, 101, 102, 103. In some studies, in mice with autoimmune disorders, the detection of fewer antibodies in those lacking the IFN-γR1 receptor suggested that there may be a positive relationship between autoantibody formation and IFN-γ. In addition, it is determined that Th1 and Th2 related Ig isotypes are equally affected 104.
IFN-γ is the most important activator of macrophages. Nitric oxide production and MHC class II regulation mechanism of macrophages are IFN-γ dependent. These two effector mechanisms are required for cell-mediated and Th1-type immune responses. IL-12, a cytokine, is released from IFN- γ activated macrophages. In addition, IL-12 directs T-helper cells to differentiation into Th1 cells 71, 105, 106. However, it is known that systemic administration of IL-12 causes a decrease in bone marrow cells, inhibiting hematopoiesis and a significant decrease in blood lymphocyte count 107. It also increases the flow of active macrophages and NK cells to the spleen.
IFN-γ secreted by Th1 cells activates phagocytic macrophages. In addition, inducible nitric oxide synthase (iNOS), which catalyzes nitrogen oxide derivative (NOx) free radicals, is activated to directly kill intracellular bacteria and protozoa (Figure 4). Mutations at IFN-γR1/2 receptors severely affect IFN-γ-mediated immunity 74, 82, 108. A mutation in the IFN- γ R chains has been found to be associated with idiopathic mycobacterial infection, and IFN-γ plays a critical role in mediating immunity against many infections such as antimycobacterial agents 109. It is considered necessary to respond to IFN-γ in the defense mediated by macrophages against intracellular pathogens. The most important function of IFN-γ, which stimulates IFN-γR, is the cellular response starting through JAK1, JAK2 and STAT1. These stimulated cells play an important role in both natural and acquired immune responses. Macrophage activation of IFN-γ is critical for its function and antigen presentation. The response that starts with IFN-γ is mandatory for host defense against many intracellular pathogens (such as mycobacterium and listeria), including viruses, bacteria and protozoa. Although it is not mandatory in host defense against some pathogens, IFN-γ has been shown to participate and contribute to the defense. It has been found that IFN- γ, like a self-feeding stimulant, stimulates NK cells and Thl cells to produce more IFN-γ by secreting IL-12 from macrophages 72, 110. It also takes part in the activation of neutrophils, which play an important role in the first line of defense.
Figure 4. Stimulation of macrophage with IFN-γ. In the stimulated macrophage, inducible nitric oxide synthase (iNOS) is activated to produce nitrogen oxide derivative (NOx) free radicals to directly kill intracellular bacteria and protozoa. Also, increased IL-12 release activates NK cells more. 
Download figure
In a study of mice with IFN-γR1-null, the immune system did not respond to IFN-γ, although they had normal leukocyte populations 111. Therefore, it was concluded that cells are deprived of the biological effects of IFN-γ, including antiviral and antitumor activity. This result was an indication that IFN-γ is important for cells to show antiviral resistance. However, even though it is so important in antiviral activation, the high pleiotropic effects of IFN-γ, which have quite wide and different effects, have limited its use as an immunotherapeutic agent to date. Nevertheless, IFN-γ continues to be tried in the treatment of SARS-coronavirus in combination with IFN α or β 112, 113. Because the need for broad-spectrum emergency treatment against microbial pathogens that still have no specific treatment remains. One of the agents that can best meet this need is thought to be IFN-γ.
Conclusion
Since IFN-γ has both natural and cellular immune system stimulation as an immunomodulating agent, has low resistance development potential and has broad spectrum antiviral properties, it should be tried alone or in combination with IFN-α-2b in broad spectrum antiviral therapy, including COVID-19, taking into account the proinflammatory/inflammatory status and timing.
References
- 1.Montecino-Rodriguez E, Berent-Maoz B, Dorshkind K. (2013) Causes, consequences, and reversal of immune system aging.https://doi.org/10.1172/JCI64096. , J Clin Invest 123, 958-965.
- 2.Thompson W W, Shay D K, Weintraub E, Brammer L, Cox N. (2003) . Mortality Associated With Influenza and Respiratory Syncytial Virus in the United States.https://doi.org/10.1001/jama.289.2.179. JAMA 289: 179.
- 4.Bleuven C, Landry C R. (2016) Molecular and cellular bases of adaptation to a changing environment in microorganisms.https://doi.org/10.1098/rspb.2016.1458. , Proc R Soc B Biol Sci 283, 20161458.
- 5.Peeri N C, Shrestha N, Rahman M S, Zaki R, Tan Z. (2020) The SARS, MERS and novel coronavirus (COVID-19) epidemics, the newest and biggest global health threats: what lessons have we learned? Int J Epidemiol pii:. dyaa033. https://doi.org/10.1093/ije/dyaa033
- 6. (2006) Immunomodulation C on ND in the S of AT. Promising Approaches to the Development of Immunomodulation for the Treatment of Infectious Diseases. , Washington (DC):NationalAcademiesPress(US)
- 7.Casadevall A, Pirofski L. (2003) The damage-response framework of microbial pathogenesis. , Nat Rev Microbiol.https://doi.org/10.1038/nrmicro732 1, 17-24.
- 8.Janeway C, Travers P, Walport M, Shlomchik M. (2001) . The Immune System in Health and Disease. Immunobiology. 5th baskı , New York:GarlandScience 425-596.
- 9.Abreu M T, Arditi M. (2004) Innate immunity and toll-like receptors: clinical implications of basic science research. , J Pediatr.https://doi.org/10.1016/j.jpeds.2004.01.057 144, 421-429.
- 11.Averdam A, Petersen B, Rosner C, Neff J, Roos C. (2009) A novel System of polymorphic and diverse NK cell receptors in primates. PLoS Genet 5,e1000688. https://doi.org/10.1371/journal.pgen.1000688
- 12.Ljunggren H-G, Kärre K. (1990) In search of the ‘missing self’: MHC molecules and NK cell recognition. , Immunol Today.https://doi.org/10.1016/0167-5699(90)90097-S 11, 237-244.
- 13.Carrillo-Bustamante P, Kesmir C, de Boer RJ. (2015) . Can Selective MHC Downregulation Explain the Specificity and Genetic Diversity of NK Cell Receptors? Front Immunol 6,311. https://doi.org/10.3389/fimmu.2015.00311 .
- 14.Kyuwa S, Cohen M, Nelson G, Tahara S M, Stohlman S A. (1994) Modulation of cellular macromolecular synthesis by coronavirus: implication for pathogenesis. , J Virol.https://doi.org/10.1128/JVI.68.10.6815-6819.1994 68, 6815-6819.
- 15.Breitbart M, Rohwer F. (2005) Here a virus, there a virus, everywhere the same virus? Trends Microbiol. 13, 278-84.
- 16.Prokunina-Olsson L, Alphonse N, Dickenson R E, Durbin J E, Glenn J S et al. (2020) COVID-19 and emerging viral infections: The case for interferon lambda. J Exp Med.https://doi.org/10.1084/jem.20200653. 217.
- 17.Ren Z, Yan L, Zhang N, Guo Y, Yang C. (2013) The newly emerged SARS-Like coronavirus HCoV-EMC also has an “Achilles’ heel”: current effective inhibitor targeting a 3C-like protease. , Protein Cell.https://doi.org/10.1007/s13238-013-2841-3 4, 248-250.
- 18.Ludwig S. (2011) Disruption of virus-host cell interactions and cell signaling pathways as an anti-viral approach against influenza virus infections. , Biol Chem.https://doi.org/10.1515/BC.2011.121 392, 837-847.
- 19.Josset L, Textoris J, Loriod B, Ferraris O, Moules V. (2010) Gene Expression Signature-Based Screening Identifies New Broadly Effective Influenza A Antivirals. PLoS One 5,e13169. https://doi.org/10.1371/journal.pone.0013169
- 20.Tisoncik J R, Korth M J, Simmons C P, Farrar J, Martin T R. (2012) Into the eye of the cytokine storm. , Microbiol Mol Biol Rev.https://doi.org/10.1128/MMBR.05015-11 76, 16-32.
- 21.Meijvis S C, Hardeman H, Remmelts H H, Heijligenberg R, Rijkers G T. (2011) Dexamethasone and length of hospital stay in patients with community-acquired pneumonia: a randomised, double-blind, placebo-controlled trial. , Lancet.https://doi.org/10.1016/S0140-6736(11)60607-7 377, 2023-2030.
- 22.Ge D, Fellay J, Thompson A J, Simon J S, Shianna K V. (2009) Genetic variation in IL28B predicts hepatitis C treatment-induced viral clearance. , Nature.https://doi.org/10.1038/nature08309 461, 399-401.
- 23.Brun-Buisson C, J-CM Richard, Mercat A, ACM Thiébaut, Brochard L. (2011) . Early Corticosteroids in Severe Influenza A/H1N1 Pneumonia and Acute Respiratory Distress Syndrome. Am J Respir Crit Care Med.https://doi.org/10.1164/rccm.201101-0135OC 183, 1200-1206.
- 24.Wang R, Jaw J J, Stutzman N C, Zou Z, Sun P D. (2012) Natural killer cell-produced IFN-γ and TNF-α induce target cell cytolysis through up-regulation of ICAM-1. , J Leukoc Biol.https://doi.org/10.1189/jlb.0611308 91, 299-309.
- 25.Polderman K H, ARJ Girbes. (2004) Drug intervention trials in sepsis: divergent results. , Lancet.https://doi.org/10.1016/S0140-6736(04)16259-4 363, 1721-1723.
- 26.Meurs E, Chong K, Galabru J, NSB Thomas, Kerr I M. (1990) Molecular cloning and characterization of the human double-stranded RNA-activated protein kinase induced by interferon. , Cell 62, 379-390.
- 27.Samuel C E. (2001) Antiviral actions of interferons. , Clin Microbiol Rev.https://doi.org/10.1128/CMR.14.4.778-809.2001 14, 778-809.
- 28.S V Kotenko, Gallagher G, V, Lewis-Antes A, Shen M et al. (2003) IFN-λs mediate antiviral protection through a distinct class II cytokine receptor complex. , Nat Immunol.https://doi.org/10.1038/ni875 4, 69-77.
- 29.Donnelly R P, S V Kotenko. (2010) Interferon-Lambda: A New Addition to an Old Family. , J Interf Cytokine Res 30, 555-64.
- 30.Lee A J, Chen B, Chew M V, Barra N G, Shenouda M M. (2017) Inflammatory monocytes require type I interferon receptor signaling to activate NK cells via IL-18 during a mucosal viral infection. , J Exp Med 214, 1153-1167.
- 31.Lee A J, Ashkar A A. (2018) The dual nature of type I and type II interferons. Front Immunol 9,2061. https://doi.org/10.3389/fimmu.2018.02061
- 32.Strander H. (1989) The action of interferons on virus-associated human neoplasms. , Cancer Surv 8-755.
- 33.Mandelli F, Avvisati G, Amadori S, Boccadoro M, Gernone A. (1990) Maintenance Treatment with Recombinant Interferon Alfa-2b in Patients with Multiple Myeloma Responding to Conventional Induction Chemotherapy. , N Engl J Med 322, 1430-1434.
- 34.Borden E C, Sen G C, Uze G, Silverman R H, Ransohoff R M. (2007) Interferons at age 50: past, current and future impact on biomedicine. Nat Rev Drug Discov 6,975-990. https://doi.org/10.1038/nrd2422
- 35.Jakimovski D, Kolb C, Ramanathan M, Zivadinov R, Weinstock-Guttman B. (2018) Interferon β for multiple sclerosis. Cold Spring Harb Perspect Med 8,a032003. https://doi.org/10.1101/cshperspect.a032003
- 36.Sheahan T P, Sims A C, Leist S R, Schäfer A, Won J. (2020) Comparative therapeutic efficacy of remdesivir and combination lopinavir, ritonavir, and interferon beta against MERS-CoV. , Nat Commun.https://doi.org/10.1038/s41467-019-13940-6 11, 222.
- 37.JF-W Chan, Yao Y, Yeung M-L, Deng W, Bao L. (2015) Treatment With Lopinavir/Ritonavir or Interferon-β1b Improves Outcome of MERS-CoV Infection in a Nonhuman Primate Model of Common Marmoset. , J Infect Dis.https://doi.org/10.1093/infdis/jiv392 212, 1904-1913.
- 38.Sallard E, Lescure F-X, Yazdanpanah Y, Mentre F, Peiffer-Smadja N. (2020) Type 1 interferons as a potential treatment against COVID-19. Antiviral Res 178,104791. https://doi.org/10.1016/j.antiviral.2020.104791
- 39.Siddiqi H K, Mehra M R. (2020) COVID-19 illness in native and immunosuppressed states: A clinical-therapeutic staging proposal. J Hear Lung Transplant. https://doi.org/10.1016/j.healun.2020.03.012
- 40.Zhou F, Yu T, Du R, Fan G, Liu Y. (2020) Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. , Lancet.https://doi.org/10.1016/S0140-6736(20)30566-3 395, 1054-1062.
- 41.Zhang W, Zhao Y, Zhang F, Wang Q, Li T. (2020) The use of anti-inflammatory drugs in the treatment of people with severe coronavirus disease 2019 (COVID-19): The Perspectives of clinical immunologists from China. Clin Immunol 214,108393. https://doi.org/10.1016/j.clim.2020.108393
- 42.Deonarain R, Alcami A, Alexiou M, Dallman M J, Gewert D R. (2000) Impaired Antiviral Response and Alpha/Beta Interferon Induction in Mice Lacking Beta Interferon. , J Virol.https://doi.org/10.1128/JVI.74.7.3404-3409.2000 74, 3404-3409.
- 43.Omrani A S, Saad M M, Baig K, Bahloul A, Abdul-Matin M. (2014) Ribavirin and interferon alfa-2a for severe Middle East respiratory syndrome coronavirus infection: a retrospective cohort study. , Lancet Infect Dis.https://doi.org/10.1016/S1473-3099(14)70920-X 14, 1090-1095.
- 44.Gao L, Yu S, Chen Q, Duan Z, Zhou J. (2010) A randomized controlled trial of low-dose recombinant human interferons α-2b nasal spray to prevent acute viral respiratory infections in military recruits. , Vaccine.https://doi.org/10.1016/j.vaccine.2010.03.062 28, 4445-4451.
- 45.Liu Y-J. (2005) IPC: Professional Type 1 Interferon-Producing Cells and Plasmacytoid Dendritic Cell Precursors. Annu Rev Immunol 23,275-306. https://doi.org/10.1146/annurev.immunol.23.021704.115633
- 46.Schneider W M, Chevillotte M D, Rice C M. (2014) Interferon-Stimulated Genes: A Complex Web of Host Defenses. , Annu Rev Immunol.https://doi.org/10.1146/annurev-immunol-032713-120231 32, 513-545.
- 47.Totura A L, Baric R S. (2012) SARS coronavirus pathogenesis: host innate immune responses and viral antagonism of interferon. , Curr Opin Virol 2, 264-275.
- 48.Billiau A. (1996) Interferon-γ: Biology and Role in Pathogenesis. , Adv. Immunol., Academic Press Inc.https://doi.org/10.1016/S0065-2776(08)60428-9 62, 61-130.
- 49.Lund F E.Randall TD.(2010) Effector and regulatory B cells: modulators of CD4+ T cell immunity. Nat Rev Immunol 10,236-247. https://doi.org/10.1038/nri2729
- 50.Baumgarth N. (2011) The double life of a B-1 cell: self-reactivity selects for protective effector functions. , Nat Rev Immunol.https://doi.org/10.1038/nri2901 11, 34-46.
- 51.Wojciechowski W, Harris D P, Sprague F, Mousseau B, Makris M. (2009) Cytokine-producing effector B cells regulate type 2 immunity to H. polygyrus. Immunity 30,421-433. https://doi.org/10.1016/j.immuni.2009.01.006
- 52.Poe J C, Smith S H, Haas K M, Yanaba K, Tsubata T. (2011) . Amplified B lymphocyte CD40 signaling drives regulatory B10 cell expansion in mice. PLoS One 6,e22464. https://doi.org/10.1371/journal.pone.0022464 .
- 53.Tu W, Lau Y-L, Zheng J, Liu Y, Chan P-L. (2008) Efficient generation of human alloantigen-specific CD4+ regulatory T cells from naive precursors by CD40-activated B cells. , Blood.https://doi.org/10.1182/blood-2008-04-152041 112, 2554-2562.
- 54.Yang M, Sun L, Wang S, Ko K-H, Xu H. (2010) Novel Function of B Cell-Activating Factor in the Induction of IL-10-Producing Regulatory B Cells. , J Immunol.https://doi.org/10.4049/jimmunol.0902551 184, 3321-3325.
- 55.Horikawa M, Minard-Colin V, Matsushita T, Tedder T F. (2011) Regulatory B cell production of IL-10 inhibits lymphoma depletion during CD20 immunotherapy in mice. , J Clin Invest 121, 4268-4280.
- 56.Bao Y, Liu X, Han C, Xu S, Xie B. (2014) Identification of IFN-γ-producing innate B cells. , Cell Res.https://doi.org/10.1038/cr.2013.155 24, 161-176.
- 57.Gray D, Gray M, Barr T. (2007) Innate responses of B cells. , Eur J Immunol.https://doi.org/10.1002/eji.200737728 37, 3304-3310.
- 58.Sindhava V J, Tuna H, Gachuki B W, DiLillo D J, Avdiushko M G. (2012) Bone Marrow Dendritic Cell-Mediated. , Regulation of TLR and B Cell Receptor Signaling in B Cells. J Immunol.https://doi.org/10.4049/jimmunol.1101352 189, 3355-3367.
- 59.Bossen C, Cachero T G, Tardivel A, Ingold K, Willen L. (2008) TACI, unlike BAFF-R, is solely activated by oligomeric BAFF and APRIL to support survival of activated B cells and plasmablasts. , Blood 111, 1004-1012.
- 60.Balázs M, Martin F, Zhou T, Kearney J F. (2002) Blood Dendritic Cells Interact with Splenic Marginal Zone B Cells to Initiate T-Independent Immune Responses. , Immunity.https://doi.org/10.1016/S1074-7613(02)00389-8 17, 341-352.
- 61.Cerutti A, Puga I, Cols M. (2011) Innate control of B cell responses. Trends Immunol 32,202-211. https://doi.org/10.1016/j.it.2011.02.004
- 62.Presti R M, Pollock J L, Dal Canto AJ, O’Guin A K, Virgin H W. (1998) . Interferon γ Regulates Acute and Latent Murine Cytomegalovirus Infection and Chronic Disease of the Great Vessels. J Exp Med 188,577-588. https://doi.org/10.1084/jem.188.3.577 .
- 63.Harris D P, Haynes L, Sayles P C, Duso D K, Eaton S M. (2000) Reciprocal regulation of polarized cytokine production by effector B and T cells. , Nat Immunol.https://doi.org/10.1038/82717 1, 475-482.
- 64.Barr T A, Brown S, Ryan G, Zhao J, Gray D. (2007) TLR-mediated stimulation of APC: Distinct cytokine responses of B cells and dendritic cells. , Eur J Immunol.https://doi.org/10.1002/eji.200636483 37, 3040-3053.
- 65.Yoshimoto T, Okamura H, Tagawa Y-I, Iwakura Y, Nakanishi K. (1997) Interleukin 18 together with interleukin 12 inhibits IgE production by induction of interferon- production from activated B cells. Proc Natl Acad Sci 94,3948-3953. https://doi.org/10.1073/pnas.94.8.3948 .
- 66.Ganapamo F, Dennis V A, Philipp M T. (2001) CD19+ cells produce IFN-γ in mice infected withBorrelia burgdorferi. , Eur J Immunol.https://doi.org/10.1002/1521-4141 31, 3460-3468.
- 67.Kubota K, Kadoya Y. (2011) Innate IFN-γ-Producing Cells in the Spleen of Mice Early after Listeria monocytogenes Infection:. Importance of Microenvironment of the Cells Involved in the Production of Innate IFN-γ. Front Immunol.https://doi.org/10.3389/fimmu.2011.00026 2-6.
- 68.Williams M A, Schmidt R L, Lenz L L. (2012) Early events regulating immunity and pathogenesis during Listeria monocytogenes infection. Trends Immunol 33,488-495. https://doi.org/10.1016/j.it.2012.04.007
- 69.Pamer E G. (2004) Immune responses to Listeria monocytogenes. , Nat Rev Immunol.https://doi.org/10.1038/nri1461 4, 812-823.
- 70.Wheelock E F. (1965) Interferon-Like Virus-Inhibitor Induced in Human Leukocytes by Phytohemagglutinin. Science.https://doi.org/10.1126/science.149.3681.310 80(149), 310-311.
- 71.Boehm U, Klamp T, Groot M, Howard J C. (1997) Cellular responses to interferon-γ. Annu Rev Immunol 15,749-95. https://doi.org/10.1146/annurev.immunol.15.1.749
- 73.Kotenko S V, Izotova L S, Pollack B P, Mariano T M, Donnelly R J. (1995) Interaction between the Components of the Interferon γ Receptor Complex. , J Biol Chem.https://doi.org/10.1074/jbc.270.36.20915 270, 20915-20921.
- 74.Farrar M A, Fernandez-Luna J, Schreiber R D. (1991) Identification of two regions within the cytoplasmic domain of the human interferon-gamma receptor required for function. , J Biol Chem 266, 19626-19635.
- 75.Zhu J, Paul W E. (2008) CD4 T cells: fates, functions, and faults. , Blood.https://doi.org/10.1182/blood-2008-05-078154 112, 1557-1569.
- 76.Rawlings J S, Rosler K M, Harrison D A. (2004) The JAK/STAT signaling pathway. , J Cell Sci.https://doi.org/10.1242/jcs.00963 117, 1281-1283.
- 77.Hemmi S, Peghini P, Metzler M, Merlin G, Dembic Z. (1989) Cloning of murine interferon gamma receptor cDNA: expression in human cells mediates high-affinity binding but is not sufficient to confer sensitivity to murine interferon gamma. Proc Natl Acad Sci 86,9901-9905. https://doi.org/10.1073/pnas.86.24.9901
- 78.Marsters S A, Pennica D, Bach E, Schreiber R D, Ashkenazi A. (1995) Interferon gamma signals via a high-affinity multisubunit receptor complex that contains two types of polypeptide chain. Proc Natl Acad Sci 92,5401-5405. https://doi.org/10.1073/pnas.92.12.5401 .
- 79.Cook J R, Jung V, Schwartz B, Wang P, Pestka S. (1992) Structural analysis of the human interferon gamma receptor: a small segment of the intracellular domain is specifically required for class I major histocompatibility complex antigen induction and antiviral activity. , Proc Natl Acad Sci 89, 11317-11321.
- 80.Farrar M A, Campbell J D, Schreiber R D. (1992) Identification of a functionally important sequence in the C terminus of the interferon-gamma receptor. Proc Natl Acad Sci 89,11706-11710. https://doi.org/10.1073/pnas.89.24.11706
- 81.Lee E-Y, KLW Schultz, Griffin D E. (2013) Mice deficient in interferon-gamma or interferon-gamma receptor 1 have distinct inflammatory responses to acute viral encephalomyelitis. PLoS One 8,e76412. https://doi.org/10.1371/journal.pone.0076412
- 82.Lu B, Ebensperger C, Dembic Z, Wang Y, Kvatyuk M. (1998) Targeted disruption of the interferon- receptor 2 gene results in severe immune defects in mice. , Proc Natl Acad Sci 95, 8233-8238.
- 83.Bazan J F. (1990) Structural design and molecular evolution of a cytokine receptor superfamily. Proc Natl Acad Sci 87,6934-6938. https://doi.org/10.1073/pnas.87.18.6934 .
- 84.Igarashi K, Garotta G, Ozmen L, Ziemiecki A, Wilks A F. (1994) Interferon-gamma induces tyrosine phosphorylation of interferon-gamma receptor and regulated association of protein tyrosine kinases. Jak1 and Jak2, with , J Biol Chem 269, 14333-14336.
- 85.Kaplan D H, Greenlund A C, Tanner J W, Shaw A S, Schreiber R D. (1996) Identification of an Interferon- Receptor Chain Sequence Required for JAK-1 Binding. , J Biol Chem.https://doi.org/10.1074/jbc.271.1.9 271, 9-12.
- 86.Greenlund A C, Farrar M A, Viviano B L, Schreiber R D. (1994) Ligand-induced IFN gamma receptor tyrosine phosphorylation couples the receptor to its signal transduction system (p91). , EMBO J.https://doi.org/10.1002/j.1460-2075.1994.tb06422.x 13, 1591-1600.
- 87.Briscoe J, Rogers N C, Witthuhn B A, Watling D, Harpur A G. (1996) Kinase-negative mutants of JAK1 can sustain interferon-gamma-inducible gene expression but not an antiviral state. , EMBO J.https://doi.org/10.1002/j.1460-2075.1996.tb00415.x 15, 799-809.
- 88.Pang Q, Fagerlie S, Christianson T A, Keeble W, Faulkner G. (2000) The fanconi anemia protein FANCC binds to and facilitates the activation of STAT1 by gamma interferon and hematopoietic growth factors. , Mol Cell Biol.https://doi.org/10.1128/MCB.20.13.4724-4735.2000 20, 4724-4735.
- 89.Heim M, Kerr I, Stark G, Darnell J. (1995) Contribution of STAT SH2 groups to specific interferon signaling by the Jak-STAT pathway. , Science.https://doi.org/10.1126/science.7871432 80(267), 1347-1349.
- 90.Schindler C, Shuai K, Prezioso V, Darnell J. (1992) Interferon-dependent tyrosine phosphorylation of a latent cytoplasmic transcription factor. , Science.https://doi.org/10.1126/science.1496401 80(257), 809-813.
- 91.Stark G R, Darnell J E. (2012) The JAK-STAT pathway at twenty. , Immunity.https://doi.org/10.1016/j.immuni.2012.03.013 36, 503-514.
- 92.Michalska A, Blaszczyk K, Wesoly J, HAR Bluyssen. (2018) A Positive Feedback Amplifier Circuit That Regulates Interferon (IFN)-Stimulated Gene Expression and Controls Type I and Type II IFN Responses. Front Immunol 9,1135. https://doi.org/10.3389/fimmu.2018.01135
- 93.Kiu H, Nicholson S E. (2012) Biology and significance of the JAK/STAT signalling pathways. , Growth Factors.https://doi.org/10.3109/08977194.2012.660936 30, 88-106.
- 94.David M, Petricoin E, Benjamin C, Pine R, Weber M. (1995) Requirement for MAP kinase (ERK2) activity in interferon alpha- and interferon beta-stimulated gene expression through STAT proteins. , Science.https://doi.org/10.1126/science.7569900 80(269), 1721-1723.
- 95.Früh K, Yang Y. (1999) Antigen presentation by MHC class I and its regulation by interferon γ. , Curr Opin Immunol.https://doi.org/10.1016/S0952-7915(99)80014-4 11, 76-81.
- 96.Cresswell P. (1994) Assembly, Transport, and Function of MHC Class II Molecules. Annu Rev Immunol 12,259-291. https://doi.org/10.1146/annurev.iy.12.040194.001355
- 97.Mond J J, Carman J, Sarma C, Ohara J, Finkelman F D. (1986) Interferon-gamma suppresses B cell stimulation factor (BSF-1) induction of class II MHC determinants on B cells. , J Immunol 137, 3534-3537.
- 98.Raval A, Puri N, Rath P C, Saxena R K. (1998) Cytokine regulation of expression of class I MHC antigens. Exp Mol Med 30,1-13. https://doi.org/10.1038/emm.1998.1
- 99.Fong T A, Mosmann T R. (1990) Alloreactive murine CD8+ T cell clones secrete the Th1 pattern of cytokines. , J Immunol 144, 1744-1752.
- 100.Maillard I, Launois P, Xenarios I, Louis J A, Acha-Orbea H. (1998) Immune Response to Mouse Mammary Tumor Virus in Mice Lacking the Alpha/Beta Interferon or the Gamma Interferon Receptor. , J Virol 72-2638.
- 101.Planz O, Ehl S, Furrer E, Horvath E, Bründler M A. (1997) A critical role for neutralizing-antibody-producing B cells, CD4+ T cells, and interferons in persistent and acute infections of mice with lymphocytic choriomeningitis virus: Implications for adoptive immunotherapy of virus carriers. Proc Natl Acad Sci USA 94,6874-6879. https://doi.org/10.1073/pnas.94.13.6874 .
- 102.Weck K E, Dal Canto AJ, Gould J D, O’Guin A K, Roth K A. (1997) Murine γ-herpesvirus 68 causes severe large-vessel arteritis in mice lacking interferon-γ responsiveness: A new model for virus-induced vascular disease. , Nat Med 3, 1346-1353.
- 103.Kang S, Brown H M, Hwang S. (2018) Direct antiviral mechanisms of interferon-gamma. Immune Netw 18,e33. https://doi.org/10.4110/in.2018.18.e33 .
- 104.Ozmen L, Roman D, Fountoulakis M, Schmid G, Ryffel B. (1995) Experimental therapy of systemic lupus erythematosus: the treatment of NZB/W mice with mouse soluble interferon-γ receptor inhibits the onset of glomerulonephritis. , Eur J Immunol.https://doi.org/10.1002/eji.1830250103 25, 6-12.
- 105.de Groen RA, Boltjes A, Hou J, Liu B-S, McPhee F. (2015) IFN-λ-mediated IL-12 production in macrophages induces IFN-γ production in human NK cells. , Eur J Immunol.https://doi.org/10.1002/eji.201444903 45, 250-259.
- 106.Yoshida A, Koide Y, Uchijima M, Yoshida T O. (1994) . IFN-γ Induces IL-12 mRNA Expression by a Murine Macrophage Cell Line, J774. Biochem Biophys Res Commun 198,857-861. https://doi.org/10.1006/bbrc.1994.1122 .
- 107.Car B D, Eng V M, Schnyder B, LeHir M, Shakhov A N. (1995) Role of interferon-γ in interleukin 12-induced pathology in mice. , Am J Pathol 147, 1693-1707.
- 108.Kotenko S V, Izotova L S, Pollack B P, Mariano T M, Donnelly R J. (1995) Interaction between the Components of the Interferon γ Receptor Complex. , J Biol Chem.https://doi.org/10.1074/jbc.270.36.20915 270, 20915-20921.
- 109.GAW Rook. (1997) Intractable mycobacterial infections associated with genetic defects in the receptor for interferon gamma: what does this tell us about immunity to mycobacteria?. , Thorax 52, 41-46.
- 110.Holland S M. (2000) Treatment of infections in the patient with Mendelian susceptibility to mycobacterial infection. , Microbes Infect 2, 1579-1590.
- 111.Lee E Y, KLW Schultz, Griffin D E. (2013) . Mice Deficient in Interferon-Gamma or Interferon-Gamma Receptor 1 Have Distinct Inflammatory Responses to Acute Viral Encephalomyelitis. PLoS One.https://doi.org/10.1371/journal.pone.0076412 8(10), 76412.
Cited by (4)
This article has been cited by 4 scholarly works according to:
Citing Articles:
Frontiers in Medicine (2023) OpenAlex Crossref Semantic Scholar
International Journal of Clinical Practice (2021) OpenAlex Crossref Semantic Scholar
Journal of Health Sciences and Medicine (2021) OpenAlex Crossref Semantic Scholar
Journal of Molecular Virology and Immunology (2020) OpenAlex Semantic Scholar