Effect of Drought and Salt Stress on Cereal Crop Plants and their Proteomic and Physiological Studies
Abstract
The photosynthetic potential and underlying internal metabolism of a plant are some of the most commonly affected physiological functions as a direct consequence of stresses due to salt and water resulting in hindering plant growth and productivity. Under the influence of such detrimental stresses, a drastic alteration in a plant's osmotic requirements, hormonal production, shedding of leaves, and closure of stomata, along with a lessening in the diffusion and transportation of CO2 and H2O are commonly seen. This review unfolds with a description of the basic methodology involved in the proteomic analysis of various proteins involved in stress response along with a brief idea on identifying and obtaining a genomic sequence for proteomic studies. It then dives deep into understanding the impact of abiotic stresses such as salinity, drought and high temperatures on cereal crops such as rice and sorghum as well as the internal dynamics of tolerance mechanism unfolding during stresses have also been described. Extensive literature describing the proteomic and physiological responses to primary and secondary effects of salt stress in cereal crops emphasizing on ROS production and apoptosis, the role of osmolytes as ROS scavengers during osmotic stress and vacuolar antiporters in ionic stress along with the responses during drought stress such as the accumulation of LEA proteins and ABA-based signaling has been reviewed and critically discussed. The study also sheds light on some experimental proteomic studies conducted on the seedlings, root tissues, and shoots of rice cultivars.
Article Information
- Received
- Accepted
- Published
Academic Editor: Furqan Kabir, Aga Khan University, Karachi, Pakistan.
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2020 Kaligotla Venkata Subrahmanya Anirudh, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Corresponding author: Rajesh K. Srivastava, Department of Biotechnology, GITAM Institute of Technology, GITAM Deemed to be University, Gandhi Nagar Campus, Rushikonda, Visakhapatnam - 530045 (A.P.), India —
Competing Interests
The authors have declared that no competing interests exist.
Funding
No specific funding statement was provided by the authors.
Data Availability
No data-availability statement was provided by the authors.
Acknowledgements
The authors acknowledge the research supported by various agencies like SERB, Department of Science and Technology, New Delhi, GoI to Dr. Nasim Akhtar (CRG/2018/000517 dated: 24.06.2019) and the University Grant Commission, New Delhi to Dr Rajesh K. Srivastava (UGC/43-64/2014(SR)/: MRP-MAJOR-BIOT-2013-13973) and Dr Nasim Akhtar (F. 42-207/2013 (SR) dated: 23.03.2013).
Citation:
Introduction
The stability of a plant mainly depends on its photosynthetic capability and overall performance and productivity. Environmental stresses caused due to salt and water play a cardinal role as they influence photosynthesis both directly or indirectly hence reducing the productivity of plants significantly. When viewed directly, salts affect the overall metabolism of the plant thus limiting the amount of fluid diffusing through the stomata and resulting in its closure. In an indirect sense, it also increases the amount of oxidative stress on the plant affecting the leaf photosynthesis 1, 2, 3. Under severe drought conditions, the soil exerts salt stress, and plants start to acclimatize themselves to the surrounding environment by shedding leaves, reducing the metabolism level in the tissues, inhibiting plant growth, and solute synthesis along with stabilizing ion transport. It has also been reported that a large alteration in the general gene expression mechanism in plants under such conditions takes place so as to cope up with the external stresses. Dehydration, osmotic and ionic factors should also be considered as a response that plants deal under saline stress. Such stresses play a major role in altering the osmotic conditions of the cells, changing the rate of production of numerous hormones from the shoot and root, and hinder the diffusion and transport of carbon dioxide and water. The metabolic and physiological changes necessary for facilitating an adaptation to either biotic or abiotic stresses are mainly due to the production of systemic signals (that is the signals of osmotic stress and also those signals resulting from the variation of metabolites and hormones such as ABA) 3, 4. It has also been proven that once the leaf starts aging or reaches a stage of senescence, the amount of salt accumulation accelerates in comparison to young leaves affecting the overall photosynthetic capability of the plant 5, 6. With the rapid increase of salt concentration of the available freshwater, the only way to ensure the survival of plants is by identifying and quantifying those genes and proteins involved in causing salt tolerance and instilling them 7.
The sugar levels present in the plant cells also play a major role in determining the photosynthetic ability of the plant. For instance, under drought conditions, the concentration of sugars tends to increase in contrast to the amount of starch, and the sugars decrease once the saturation for drought stress is reached by the plant. The concentration of sugars during drought stress is important because the entire gene alteration and changes in the proteomic features of genes involved in photosynthesis are dependent on it 8, 9. The process is being studied by isolation, purification, and identification of proteome changes taking place for the tolerance of these stresses. A brief methodology involved in the proteome analysis is presented in Figure 1.
Figure 1. Methodology of Proteomic analysis.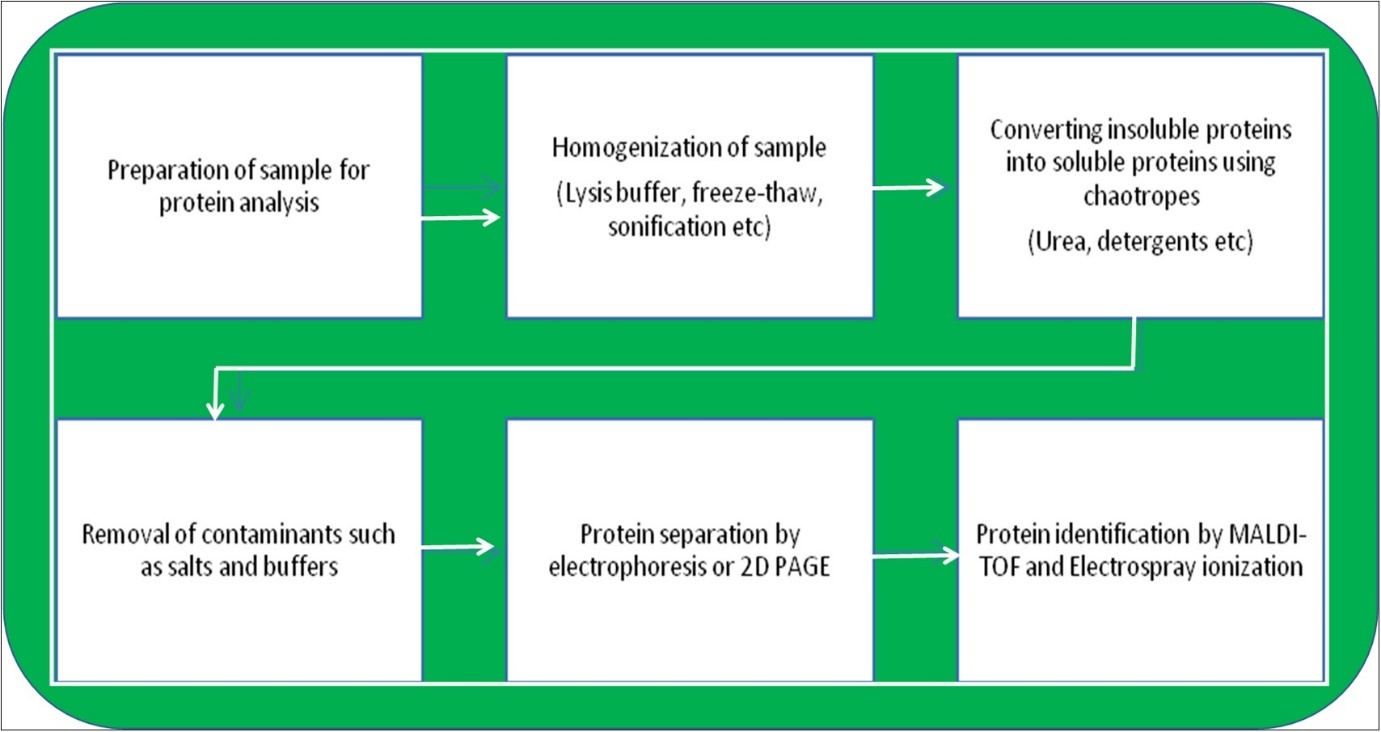
Download figure
Impact of Abiotic Stresses on Cereal Plants
Various abiotic stresses in the environment such as high salt concentration, scarcity of water, and high temperatures cause an overall decrease in the productivity and development of plants leading to death in some cases. Abiotic stresses in agricultural crops result in dwindling food supplies due to a reduction of crop yield potential by a margin greater than 50% 10, 11. Plants in semi-arid regions have increased drought and salinity stresses due to inadequate rains making irrigation their only source for survival. It is inferred from previous studies that as the severity of the drought stress increases, the number of affected genes and proteins rapidly scale 12, 13.
Analyzing the gene at the protein level quantitatively enables expression profiling thus determining their response to the plant stress which is a major step. There is a need to understand more about plant tolerance mechanisms. As there are variations in these stress responses by different cops. For instance, sorghum is much more resistant to these stresses than maize. Similarly, there is a change in the transcription profile of rice and Arabidopsis thaliana under various drought and salt stresses 14, 15. An outline of internal tolerance or resistance mechanisms by plants is presented in Figure 2.
Figure 2. Internal mechanism of Tolerance/Resistance to various stresses
Download figure
Plants react differently to the salt stress conditions by removing excess salt from the cells thus reducing its overall concentration and thereby entirely preventing its entry into the plant. Lowering the concentration of various organic acids compensate for the reduced salt levels in the cells. 16, 17. It was found that the drought and salt stress affects a large number of genes involved in the process of photosynthesis with the salt stress playing a greater role in affecting those genes in comparison to drought stress. When the plants face salt stress, the ion concentrations of sodium, potassium, and chloride ions burgeon rapidly enforcing the plants to reduce their concentration. Developing food crops adapted to such stresses is concurrent with changes in the climate and population sizes 18, 19, 20. This knowledge leads to improved food security and stress tolerance of agricultural crops by carrying out various genetic advancements.
Salt Stress
Electrical conductivity tests decipher the concentration of salts in a solution which is measured at 25 degrees Celsius with the least concentrated saline soils having an electrical conductivity value of 4 Deci Siemens/m 21. The age of the plant and the type of the species determine the impact of these abiotic stresses. As described in the recent experimental studies that plants in its germination stage is much more resistant than the latter stages of growth to salt stress and are thus classified into halophytes or glycophytes based on their sensitivity 22, 23.
Disruption of the ionic equilibrium resulting in the reactive oxygen species production which in turn reduces the protein content of seeds and overall dry weight of the plant shoot under high salt stress severely affecting the human food and animal feeds 24, 25. The proteomic studies carried out during salt stress in rice, wheat, grapevine, soybean, and potato showed a surge in the expression of oxidative stress-related proteins. Reproduction in plants like Arabidopsis was affected as the flowers being highly sensitive to ROS could not produce the minimal quantity of seeds essential for its propagation. But artificially introducing an osmolyte like glycine betaine into Arabidopsis prevents the production of malformed flowers by hindering the formation of surplus ROS 26, 27, 28. Mitochondria the powerhouse of the cell largely accounts for the production of ROS apart from undergoing substantial changes in its proteome during salt stress 29, 30. As previously reported in rice, apoptosis occurred in those cells subjected to salt stress. Glycoside hydrolase alongside other proteins was reported to play a role in initializing programmed cell death and ROS production 31.
Tremendous progress has been made in analyzing the proteome of various subcellular components such as plasma membranes, Golgi membranes, mitochondria, chloroplasts, etc, and advancement in the methods of their isolation. Among these, plasma membranes are pivotal as they act as pathways for transducing the generated stress signals 32, 33. In a study, the plasma membrane of IR651, a variety of rice having a high tolerance to salt stress was subjected to proteomic analysis which resulted in the identification of 24 proteins involved in the formation of signaling centers through efficient PPIs and their significant role in establishing a K+/Na+ balance under such conditions 34. Hence, proteomics has paved the way for understanding and elucidating the intricate expression mechanisms of proteins when subjected to salt and drought stresses.
Physiological Response to Primary and Secondary Effects of Salt Stress
Osmotic Stress
Fundamentally, osmotic stress is caused when the concentration of soluble salts in soils drastically advances resulting in a substantial decrease in the osmotic potential of the soil. This leads to a lowering of turgor pressure due to loss of water from the roots of the plants to the surroundings affecting cell division and proliferation. To combat such water losses, plants osmotically adjust the water potential in cells. By accumulating the required concentration of ions and organic solutes, plants can adjust their osmotic potential in saline conditions. For instance, sodium is utilized as an osmoticum and stored in vacuoles as they constitute a major volume of the cell storing resources and waste products whereas potassium is retained in the cytoplasm in cereal crops. Such systematic segregation of ions from the cytoplasm maintains an osmotic equilibrium along with preventing the toxic effects of these ions on cellular and metabolic processes 35, 36.
Plants tend to accumulate osmoprotectants that are basically highly soluble non-toxic and low molecular weight compatible osmolytes not in the vacuoles, but in the cytoplasm, lumen, and matrix of various cell organelles as a form of osmotic adjustment. Some of the highly prevalent osmolytes occurring in a majority of the plants are glycine betaine, proline, sugars, etc. Some exceptions like Arabidopsis and Tobacco do not contain glycine betaine and mannitol’s 37, 38. The embryos of sorghum in saline environments employ an adaptive strategy limiting their germination by accumulating larger quantities of proline. These compatible osmolytes generally function by adjusting the cells as an osmoticum and lessening the osmotic potential or for protecting various molecules inside a cell as osmoprotectants 39. Some of the common osmoprotectants like glycine betaine stabilizes various macromolecules in the cell with mannitol acting as a ROS scavenger preventing cell damage 40.
The conformation and charge interactions of the hydration layers circumscribing a protein are modified causing the protein to denature due to the presence of higher concentrated solutes like NaCl and MgSO4 in the cytoplasm. But osmoprotectants being strong structural formers with water prevents such protein unfolding’s 41, 42. Enhancing these natural osmoprotectants in crop plants in response to various abiotic stresses would improve food production from such degraded lands and it can be seen in Table 1.
Table 1. Influences of stress conditions on plant physiology with good survival| Type of stress | Effect on plants physiology | References |
|---|---|---|
| Five osmotic stress levels (0, -0.3, -0.6, -0.9, and -1.2 MPa) and five salinity stress levels (0, 3, 6, 9, and 12 dS m−1), respectively were used on plants | The highest tolerance to salinity and osmotic stresses was observed in Ghamsar-Barzak ecotype that obtained its seed was from a dry climate with the lowest mean annual precipitation and soil pH and EC are high | 68 |
| Salinity, osmotic and ABA treatment is induced the CaWRKY27 genes. CaWRKY27 functions in the response to salinity and osmotic stress | CaWRKY27 positively regulates resistance to the pathogenic bacterium Ralstonia solanacearum and negatively regulates thermotolerance. It increased sensitivity to salinity and osmotic stress, with a higher inhibition of both root elongation and whole plant growth. | 69 |
| Identification of MdTAT2 is conferred the tolerance to drought and osmotic stresses in plants. MdTATs showed distinct expression patterns in different apple tissues or under drought stress | MdTAT2 overexpression increased resistance to drought and osmotic stress. It is found Tyrosine amino transferase (TAT) genes in apple (Malus domestica). | 70 |
| Mechanisms of toxicity of imidazolium-based ionic liquids against Arabidopsis thaliana. Oxidative stress response and effects on photosynthesis of A. thaliana in ILs treatment. | Antioxidase activity (SOD, CAT, POD and GPX) changed response to ILs stress. Next ROS and MDA increased with higher ILs concentration and longer carbon chain length. It shown from biological process, cellular component, and molecular function categories. | 71 |
| 24-Epibrassinolide an active brassinolide and its role found in salt stress tolerance in plants. EBL improved photosynthesis by protecting chloroplast ultra-structure and by improving chlorophyll contents under salt stress | 24-Epibrassinolide (EBL) important brassinosteroid, played very important role in plant metabolism. EBL increases proline accumulation and other hormones under salt stress. EBL plays a crucial role as in regulating ionic homeostasis under salt stress. | 72 |
Ionic Stress
Saline stress is basically due to the presence of one of the most common salts, sodium chloride, disrupting the ionic balance of Na+ and Cl- hindering the metabolic processes of various organelles in a cell along with K+ and Ca2+. Different plant species with diverse cell types have evolved various tolerance mechanism towards ionic stresses such as excluding sodium and chloride ions from the shoots, controlling the uptake of ions from the soil, and releasing out through the leaves thus increasing the tolerance levels of these in the cytoplasm of cells 43, 44.
In the case of plants highly sensitive to saline conditions, the first possible course of action to avoid ion toxicity is to initially limit the entry of salts into the plants via their roots as seen in common bean where the absorbed sodium ions are prevented from translocating to the shoot system by a regulatory mechanism 45. Further, leaves tend to experience greater Na+ ion toxicity since the roots transport them unidirectionally through the xylem with their quantity depending on the rate of transpiration and ion concentration in the xylem vessels. To prevent the Na+ accumulation and toxicity in NaCl stress facing cell's cytoplasm, organelles, and vacuoles. The vacuolar Na+/H+ antiporters utilize the pH gradient generated through different ATPase and PPase for avoiding cytosol toxicity 46.
As the concentration of ions such as sodium, potassium increases, the water potential gradually diminishes resulting in plants imbibing low amounts of water thus modifying their whole physiological and metabolic conditions. As reported in the work done by Pang et al. 47, an increase in the biosynthesis of proteins, activity of RUBISCO, and carbon assimilation in the plant Thellungiella halophila rose to lead to the creation of a new acclimatized environment for plants under extreme salt stress conditions 47.
Plants growing in saline soils tend to spend a lot of energy for synthesizing organic osmolytes for osmoregulation along with maintaining and stabilizing the protein function 48. There exists a variation among the two types of halophytes based on the types of ions that they uptake from saline soils. The dicotyledonous variants generally tend to take up Na+ and Cl- whereas the monocotyledonous ones accumulate K+ rather than Na+. Some halophytes are equipped with salt glands or bladders in their leaves for the purpose of excreting excess Na+ ions 49, 50.
Nutrient Deficiency
A major difficulty of plants in highly saline environments is the imbibing essential ions from the soil due to the over- dominance of certain non-essential ions making them nutrient deficient. Apart from salt stress creating competition for ion uptake from the soil, it also results in stunted growth of the plants due to the development of roots with a length shorter than the average thus limiting the uptake of nutrients. The interference of Na+ ions with the K+ ion channel transporters in the roots leads to its deficiency in the developing plant cells, affecting the ongoing enzyme activity along with limiting the synthesis of proteins as it acts as a co-factor for enzyme activation along with kickstarting the binding of tRNA to ribosomes 49, 51.
Oxidative Stress
Plants when exposed to biotic and abiotic environmental stresses, their rate of production of reactive oxygen species tends to increase substantially when compared to its production from a cellular phenomenon like photorespiration and fatty acid oxidation. Other sources of ROS generation that subjects the plant cells to oxidative stress are H2O2, HO- etc. Plants employ antioxidants such as glutathione, thioredoxin, etc, and scavenging enzymes like SOD, peroxidases, and catalases for preventing damage to membrane lipids, DNA, photosystem complexes, and proteins. Some of the major cellular structures possessing considerable portions of polyunsaturated lipids inclined for damage by oxidative stress are photosynthetic cells like chloroplasts 24, 30, 52.
Drought Tolerance in Cereal Crops
The plant cells exploit various cellular, molecular, and biochemical mechanisms to keep up and maintain their normal physiological functions under water stress due to drought. One of the major challenges is to improve crop yield under drought conditions. The cellular processes in plants such as photosynthesis get largely affected leading to a collapse in its carbon dioxide assimilation rate, ultimately causing a stunted growth of the plant and a reduced crop yield. During such conditions, cells tend to lose their turgor pressure along with facing a considerable increase in the concentration of solutes. In order to cope up with such losses of turgor pressure, plants osmotically adjust it with compatible solutes like glycine betaine and sugars as can be seen in some sorghum varieties 35, 53.
Vegetative tissues of plants facing water stress have been observed to accumulate late embryogenesis associated (LEA) proteins which help in guarding cell structures against dehydration by confiscating ions, sustaining the structures of membrane proteins, renaturing the unfolded proteins, and also operating as a hydrating buffer 54, 55. Under such conditions, a substantial rise in the antioxidant compounds and enzymes can be seen as they prevent the damage of proteins and DNA caused by oxidative stress. Along with this, protease production takes place for protein degradation and recycling to prevent the aggregation of damaged cellular structures and for renewing the synthesis of proteins 56, 57.
Proteomics Analysis of Cereal Crops Under Drought Stress
One major feature that can be seen in plants facing drought stress is the closure of stomatal pores aided by guard cells. Abscisic acid (ABA), a plant growth hormone controls this process along with producing many transcriptional factors. A guard cell proteome that was subjected to in-silico based analysis revealed 336 proteins previously unknown along with 52 signaling proteins involved in the mechanism. One such identified protein is known as myrosinase TGG1, involved in limiting the ABA-based signaling process that causes the stomata to open under drought conditions 58, 59. Proteomic studies prove to be useful for uncovering the proteins involved in novel signaling mechanisms under such drought conditions. Further, it is also understood that though there is no change in the gene expression during drought and salt stress, there may be some alterations in the protein concentration 60.
Apart from pathways that depend on ABA for acquiring tolerance during drought stress, certain genes like DREB follow an ABA independent pathway. DREB2A, a member of the DREB family and vital for multiple signaling pathways, when over-expressed, advanced the drought tolerance levels in Arabidopsis 61. Drought resistant plants could be developed by altering the pathways of ABA signaling utilizing molecules such as Sphingosine-1-phosphate as observed in Arabidopsis. Proteome research could be seen to be extremely beneficial for plant improvement if various stress-related genes and other resources could be employed in such studies 62.
Plant Proteome Studies During salt and Drought Stress
Fundamentally, plant response to stresses is relatively complicated stemming from the perception of a signal to its transduction and gene expression thus conferring tolerance to the stress. Advancements in proteomic technologies have led researchers to identify the presence of shared sensory mechanisms during stress commonly called cross-talk signaling systems. Such signaling systems are evident in plants during salt and drought stress in the form of osmotic adjustments 61. An array of experimental designs utilizing a variety of plant species, organs, and organelles, response levels to stresses, and variable sampling times has been incorporated for perceiving the spatial and temporal expression proteomics during stress and elucidated below in Figure 363.
Figure 3. Stress response-based organ specific proteomics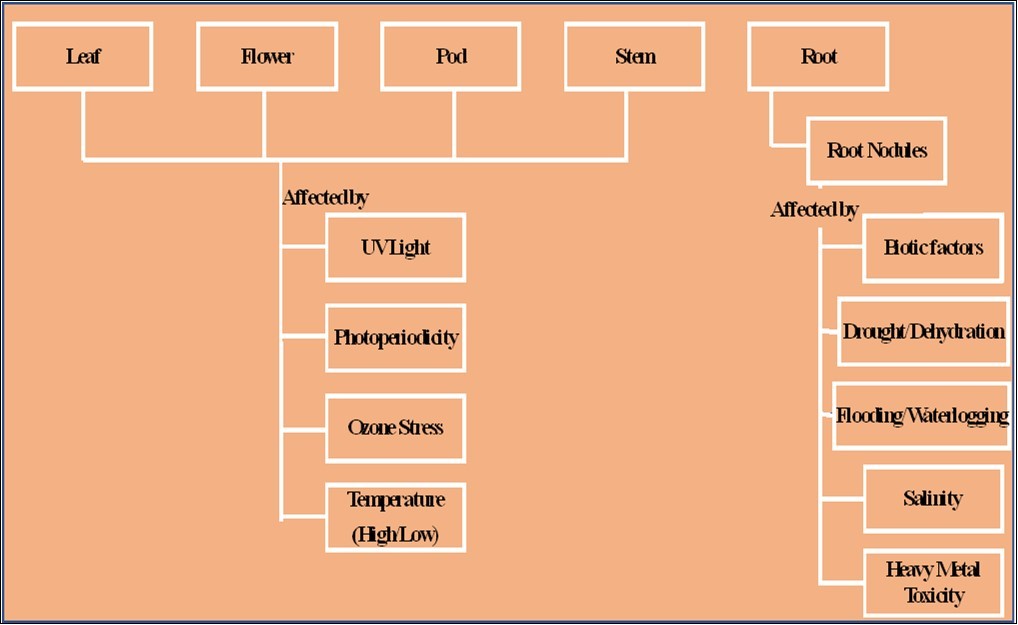
Download figure
Chourey et.al. Identified six stress proteins that arrested the growth of the seedlings among which four of them were classified as LEA proteins that were degraded during salt stress recovery studies conducted on rice 64. Zang et. al., reported the expression of 327 protein spots when the leaf sheath of 14 days old rice seedlings were subjected to osmotic stress 65. Among these, around 12 spots were found to be upregulated which constituted GST and glyoxalase. The 3 other downregulated proteins were found to be chaperone proteins. But the reason, why only those 3 proteins were downregulated could not be understood 65. In a similar study reported by Chitteti et. al., the emphasis was made on the changes in the expression of phosphoproteomes in rice roots during salt stress utilizing a fluorescent Pro-Q diamond stain. They identified 17 out of the 20 proteins that were upregulated such as GST and Hsp70. The 11 out of 18 downregulated proteins included ATP Synthase, protein kinase, etc 66. As previously discussed, subjecting plants to salt stress results in both ionic and osmotic effects characterized by a growth phase governed by 2 phases. In the first phase, the plant growth is stunted caused by osmotic stress, and if the salinity level advances, ion toxicity, and death occur due to the concentration of ions in the plant shoots 10.
According to the study performed by Salekdeh et. al., two cultivars of rice plants, one resistant and the other sensitive to stress were propagated through hydroponics where concentrations of 50mM and 100mM NaCl was introduced for a period of 2 and 3 weeks respectively 67. The root tissues and their proteins were extracted after 4 weeks and further subjected to two-dimension electrophoresis (2DE) for identifying protein expression changes. Very few spots were found to exhibit qualitative changes whereas the remaining 5 spots showed quantitative changes and the 3 proteins identified and can be seen in Figure 4.
Figure 4. Different stress condition on Plant growth and its development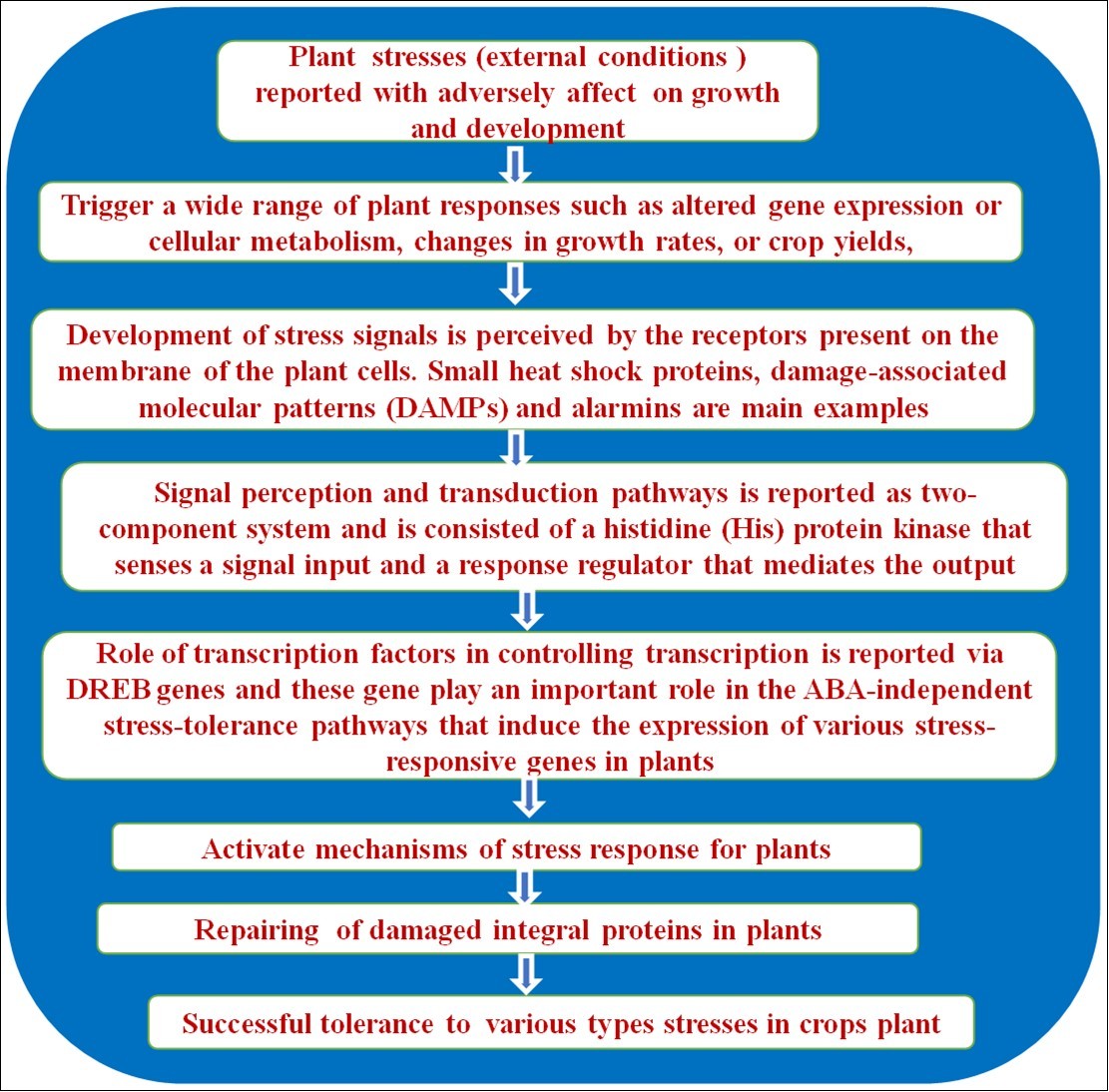
Download figure
Upon subsequent analysis with MALDI-TOF MS and ESI-Q MS/MS among them were CCOMT, ascorbate peroxidase, an ROS scavenging enzyme, and a highly stress-responsive DNA protecting ASR1-like protein were identified. They found both the cultivars had similar mechanisms for dealing with oxidative stresses except for Caffeoyl- CoA O-methyltransferase, a protein crucial for hindering the movement of sodium-rich water from the surrounding medium to the plant cell, a phenomenon termed as lignification, in the salt-resistant rice cultivar. By genetically engineering the salt stress proteins, they could be utilized as valuable molecular markers for developing highly efficient salt-tolerant crops 67.
Posttranslational Modification in Plants due to Stresses Response
Stress-induced adaptive responses are reported efficient way in the plants that depend on intricate coordination among the multiple signaling pathways. This has act in coordinate manners but in some cases, it is found in antagonistic manner. Modification in protein post-translation process is reported to regulate the protein activity and it is reported to localize with protein- interactions in various cellular processes. That can lead to elaborate the regulation of plant responses due to various external stimuli 73. Via understanding the responses among the crop plants under various field condition and that is crucial to design novel stress-tolerant cultivars and it has maintained in robust ways homeostasis under extreme conditions or factors. The proteomic studies or reports on protein post-translation modification (PTM) for different crops were discussed. Role of crop PTM is found in regulation of stress response mechanism and it is difficult to notice due to several novel factor or insights. The technique for detection of PTM in different plants under abiotic stress conditions and PTM control function is reported un representative proteins. Response of PTMs under different abiotic condition is reported soybean crops under submergence conditions. Current findings for PTMs in soybean protein is reported under flooding stresses and helped in providing information on advanced form in PTM processes in relation to plant adaptation due to abiotic stresses. The important of PTM study can provide the adequate solution in agriculture production in coming periods 73, 74. The many crop plant under natural conditions or factors is reported to various abiotic components of environment stresses and these are reported as heat wave or drought conditions via becoming more prevalent in coming years or decades. Plant acclimation as well as tolerance approaches to th abiotics stresses are reported to associate with modified PTM processes with specific proteins and is reported to important for regulation of protein functions, sub-cellular locations, stability as well as protein activity in many crops including soybean 75. The reports on plant responses to various abiotics stresses at PTMs level is reported to crucial for plant phenotypes for crops improvements. The identification and quantification of ability of PTM on a large scale can contribute to details protein functional characterization approach that can improve our understanding the processes of various crops under stress acclimation and stress tolerance conditions. There are hundreds of PTMs that can help to determine the possible protein modification mechanisms. Various types of PTMs is reported to characterize with detection approaches that can help in advances in PTMs processes with crop proteomics information. This specific PTMs in crop response to abiotics stresses is found to helpful solutions 76. The plant responses due to delicate force signal such as light tough, and are found to very similar to animal or human neural system. The demonstration of thigmotropism, thigmonastic movement or thgmomorphogenesis is reported in various crops plants. The approaches of force-signaling network can be understand via application stable isotope labeling in Arabidopsis (SILIA) tool and it can be applied for proteomic of quantitative post-translation modification that can access the protein phosphorylation medications in Arabidopsis species 77. This plant species is subjected to nearly forty second cotton-swab toruc experiments and these have identified a 4895 non-redundant phosphopeptides and 579 unreported phosphosites, 509 phosphoproteins as well as 24 touch regulated phosphoprotein (TREPH) groups. Further, molecular biological, genetics or bioinformatics analyses can also report the uncharacterized TREPH1 protein; require for bottling-delay regards in Arabidopsis species due to touch response. Protein phosphorylation and TREPH1 protein are found for mechanotransduction pathway due to plant thgmomorphogenesis 78.
Phosphorylation Processes in Signal Transduction in plants due to Stresses Conditions Response
Various types of stress-signaling pathways are reported to play crucial role in the maintenance of homoeostasis in plant cells and these pathways is shown important role adaptation of new cellular adaptation situations. Plant organelles such as ER (endoplasmic reticulum) are reported with unfolded protein responses (UPR) with activation of biosynthetic stresses that leads to compensatory increase in ER activity. Further, The JNK (Jun N-terminal kinase) and p38 MAPK (mitogen-actvated protein kinase) signal pathway are reported to control the adaptive responses to intracellular or extracellular stresses 79. Various environmental changes i.e. U.V. light, heat or hyperosmotics conditions as well as exposure to inflammatory cytokines are reported. Metabolic stress due to high-fat diet can be good stimulus with coordination of activities from UPR and JNK/38 signaling pathways. Chronic activation due to stress responses pathways can cause metabolic changes with association with obesity and altered insulin sensitive capability 1, 2. ER homoeostasis is regulated by network of signal pathways due to stearoyl-coA desaturase (SCD), p-38 MAPK and also UPR responses. All these pathways are found to locate at the interface of cell cycle control and cell stress. Cross-regulation can also help to various plant stresses. Interference with SCD-1 with small interfering (si) RNA or the specific SCD-1 inhibitors can induced phosphorylation with activation of p38 MARK in NEH-313 mouse fibroblast 80.
There are various stresses including salt or drought signal transduction that are reported to consist of ionic or osmotic homeostasis signal pathways. Further, detoxification processes (such as damage control and repair mechanism) pathways is reported for growth regulation. Salt stress due to ionic concentration is reported to signaling via SOS (as global response to plant DNA damaging pathways) pathway and also calcium-responsive SOS3-SOS2 protein kinases complexes that can control the expression and activity of ion transporter such as SOS1. Next, osmotic stress is found to activate the several protein kinases (i.e. MAK-mitogen-actvated kinase) with mediation of osmotic homeostasis and or detoxification responses 81. There are numbers of phospholipids system that can be activated via osmotic stresses with generation of diverse arrays of messenger molecules. There is discussion on functional upstream of the osmotic stress-activated protein kinases. Abscisic acid (ABA) biosynthesis regulation is reported by osmotic stress at multiples steps. Further ABA dependent or independent osmotic stress signaling can show the modification of constitutive manner expressed transcription factors via leading to the expression of early response transcriptional activators with activation of downstream stress tolerance effectors genes 81, 82. Salt stress is reported as major environmental factors that limit plant growth and productivity. Mechanism mediating salt resistance can be understand with design ways for improving crops performance under adverse environmental condition such as salt or drought stresses. Salt stress can generate ionic stress, osmotic or secondary stresses with also oxidative stresses in various plants. Adaptation due salt stresses, plant can rely on signals and pathways via re-establishing cellular ionic, osmotic or reactive oxygen species (ROS) homeostasis. Genetic and biochemical analyses can reveal for several core stress signal pathways with participation in salt resistance 83. Salt sensitive signaling pathways can play key role in maintaining the ionic homeostasis via extruding sodium ions into the apoplast. Further, mitogen activated protein kinase (MAPK) cascade system is reported for mediation of ionic, osmotic as well as ROR homeostasis and SnRK2 (responsible for encoding 1-related protein kinase 2) protein is involved in maintaining of osmotic homeostasis. Next, identification of components and pathways, involved in plants responses to salt stress with their regulatory mechanism via also identification of sensors that involved in salt induced stress signaling pathways in plants 83, 84.
Conclusion and Prospects
Abiotic stresses such as drought, salinity, mineral and nutrient deficiency, and consequent oxidative and osmotic stress result in a radical crop yield reduction to a level greater than 50% impacting every nation trying to cope with the ever-increasing food demand. Such stresses are found to adversely affect plants growth and crop yield by hindering their ability to perform photosynthesis, cell division and proliferation and a decrease in solute synthesis and ion transport, leaf shedding, and secondary metabolite production apart from generating harmful proteins like glycoside hydrolase causing apoptosis and reactive oxygen species formation, protein unfolding’s, loosening of cell walls, proteolysis, etc.
In general, such abiotic stresses cause a substantial change in the transcription profiles of various crops and through comparative proteomics, it has been experimentally shown that salt stress affected the plant genes more than drought stress. It has also been shown that the impact varies based on the age and type of species with a plant's germination stage most likely to be heavily impacted. Similarly, studies on the role of abiotic stress in influencing the over or underproduction of several antioxidants have been carried out but the proteins involved and their characterization is yet to be known. Significant progress has been made in countering the detrimental effects of ROS during salt stress as seen in Arabidopsis were artificially induced Glycine betaine prevented the malformation of flowers.
Metabolic activities of plants heavily depend on the type of nutrients taken and scarcity in any of those may result in the generation of various signaling pathways. Thus, correlating the pathways with the expressed proteins help serve as potential indicators during mineral deficiency studies in plants. Under severe levels of water stress, a majority of the proteins expressed fall under the category of LEA proteins which help plant cells by renaturing the unfolded proteins and preventing dehydration from cellular components.
The entirety of the human and livestock population depends on cereals such as wheat, sorghum, maize, etc which constitutes nearly 92% of all the crops produced globally as the significant food for sustenance. One significant void that can be overcome by utilizing proteomic analysis over transcriptomic and genomic technologies is its ability to pave a path for understanding the intricate mechanisms related to salt and drought stress in plants, hence giving an edge in potential marker-assisted breeding methods shortly. Proteomic profiles that are of utmost importance in crop proteomic analysis include regulatory modifications, protein-protein interactions, and quantitative profiles with Gel-based and Gel-free quantitative approaches based on subcellular proteomics being widely used. Due to its immense potential in contributing knowledge at the molecular level on the role of post-translational modifications, protein-protein interactions, etc, proteomics stands out as one of the indispensable resources for characterizing plant phenotypes and breeding behavior.
Intense research is currently underway for comprehending the innate physiological and biochemical characteristics of plants during severe salt and drought stress and one way is by mapping the proteome of pre-existing protein reference maps of cereal crop plants. A possible means for understanding the various inner mechanisms of plants involved in drought stress in the future is by blending potential approaches such as transcriptomics, proteomics, genomics, and other analyses at the molecular level.
Authors’ Contributions
The design concept of the whole manuscript, literature survey and preparation were performed by KVS Anirudh under the guidance of N Akhtar. The manuscript was redesigned and rewritten by T Chakraborty. Further, N Akhtar and RK Srivastava have edited and extensively modified the manuscript. All authors read and approved the final manuscript.
Compliance with Ethical Standard
Ethical Approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Informed Consent
Informed consent is not required from all individual participants included in the study.
References
- 1.Ashraf M, Harris P J C. (2013) Photosynthesis under stressful environments: An overview. , Photosynthetica 51, 163-190.
- 2.K A Fayez, S A Bazaid. (2014) Improving drought and salinity tolerance in barley by application of salicylic acid and potassium nitrate. doi: 10.1016/j.jssas.2013.01.001Journal of the Saudi. , Society of Agricultural Sciences 13(1), 45-55.
- 3.Ali Q, Ashraf M. (2011) Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of trehalose: growth, photosynthesis, water relations and oxidative defence mechanism. doi: 10.1111/j.1439-037X.2010.00463.x.Journal of Agronomy and Crop Science,197(4):. 258-271.
- 4.Hasanuzzaman M, Nahar K, M, Roychowdhury R, Fujita M. (2013) Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. doi: 10.3390/ijms14059643.International journal of molecular sciences. 14(5), 9643-9684.
- 5.E L Devi, Kumar S, T B Singh, S K, Beemrote A. (2017) Adaptation strategies and defence mechanisms of plants during environmental stress. doi: 10.1007/978-3-319-68717-9_20. In Medicinal Plants and Environmental Challenges. , Cham 359-413.
- 6.J R Acosta-Motos, M F Ortuño, Bernal-Vicente A, Diaz-Vivancos P, M J Sanchez-Blanco.et al (2017). Plant responses to salt stress: adaptive mechanisms. doi: 10.3390/agronomy7010018. , Agronomy 7(1), 18.
- 7.G M Ali, Komatsu S. (2006) Proteomic analysis of rice leaf sheath during drought stress. doi: 10.1021/pr050291g. , Journal of proteome research 5(2), 396-403.
- 8.M, Flexas J, Pinheiro C. (2009) Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. doi: 10.1093/aob/mcn125.Annals of botany. 103(4), 551-560.
- 9.Stitt M, Gibon Y, Lunn J E, Piques M. (2007) Multilevel genomics analysis of carbon signalling during low carbon availability: coordinating the supply and utilisation of carbon in a fluctuating environment. doi: 10.1071/FP06249. , Functional Plant Biology 34, 526-549.
- 10.Gupta B, Huang B. (2014) Mechanism of salinity tolerance in plants: physiological, bio chemical, and molecular characterization. doi: 10.1155/2014/701596.International journal of genomics.
- 11.Oliveira A B de, Alencar N L M, Gomes-Filho E. (2013) Comparison between the water and salt stress effects on plant growth and development. doi: 10.5772/54223.Responses of organisms to water stress. 67.
- 12.Shrivastava P, Kumar R. (2015) Soil salinity: a serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. doi: 10.1016/j.sjbs.2014.12.001. Saudi journal of biological sciences. 22(2), 123-131.
- 13.Alqurashi M, Chiapello M, Bianchet C, Paolocci F, Lilley.K.S.et al (2018). Early responses to severe drought stress in the Arabidopsis thaliana cell suspension culture proteome. doi: 10.3390/proteomes6040038.Proteomes. 6(4), 38.
- 14.Parker R, T J Flowers, A L Moore, N V Harpham. (2006) An accurate and reproducible method for proteome profiling of the effects of salt stress in the rice leaf lamina. doi: 10.1093/jxb/erj134. , Journal of Experimental Botany 57(5), 1109-1118.
- 15.M A Omidbakhshfard, Omranian N, F S Ahmadi, Nikoloski Z, Mueller-Roeber B. (2012) Effect of salt stress on genes encoding translation-associated proteins. in Arabidopsis thaliana. doi: 10.4161/psb.21218. Plantsignaling&behavior 7(9), 1095-1102.
- 16.Carillo P, M G Annunziata, Pontecorvo G, Fuggi A, Woodrow P. (2011) Salinity stress and salt tolerance.doi: 10.5772/22331.Abiotic stress in plants–Mechanisms and adaptations,1:. 21-38.
- 17.Xie E, Wei X, Ding A, Zheng L, Wu X. (2020) . Short-Term Effects of Salt Stress on the Amino Acids of Phragmites australis Root Exudates in Constructed Wetlands. doi: 10.3390/w12020569. Water 12(2), 569.
- 18.Chaves M M, Flexas J, Pinheiro C. (2009) Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. doi: 10.1093/aob/mcn125. Annals of botany. 103(4), 551-560.
- 19.M S Khan, M N Khan. (2019) Understanding Plant Responses to Drought and Salt Stresses: Advances and Challenges in “Omics” Approaches. doi: 10.5772/intechopen.81041. In Transgenic Crops-Emerging Trends and FuturePerspectives.IntechOpen.
- 20.Chen H, J G. (2010) Osmotic adjustment and plant adaptation to environmental changes related to drought and salinity. , Environmental Reviews 18, 309-319.
- 21.S A, Zaman M, Heng L. (2018) Introduction to soil salinity, sodicity and diagnostics techniques. doi: 10.1007/978-3-319-96190-3_1. In Guideline forsalinity assessment, mitigation and adaptation using nuclear and related techniques. , Cham 1-42.
- 22.Yuan F, Guo J, Shabala S, Wang B. (2019) Reproductive physiology of halophytes: current standing. doi: 10.3389/fpls.2018.01954. Frontiers in plant science, 9.
- 23.Bolton A, Simon P. (2019) Variation for salinity tolerance during seed germination in diverse carrot [Daucus carota (L.)] germplasm. doi: 10.21273/HORTSCI13333-18. , HortScience 54(1), 38-44.
- 24.Sharma P, A B Jha, R S Dubey, Pessarakli M. (2012) Reactive oxygen species, oxidative damage, and antioxidative defence mechanism in plants under stressful conditions. doi: 10.1155/2012/217037. Journal of botany.
- 25.M I Hussain, D A Lyra, Farooq M, Nikoloudakis N, Khalid N. (2016) Salt and drought stresses in safflower: a review. doi: 10.1007/s13593-015-0344-8.Agronomy for sustainable development. 36(1), 4.
- 26.Mousavi S, Regni L, Bocchini M, Mariotti R, N G Cultrera. (2019) Physiological, epigenetic and genetic regulation in some olive cultivars under salt stress. doi: 10.1038/s41598-018-37496-5. Scientific reports. 9(1), 1-17.
- 27.Manishankar P, Wang N, Köster P, A, Kudla J. (2018) Calcium signaling during salt stress and in the regulation of ion homeostasis. doi: 10.1093/jxb/ery201. , Journal of experimental botany 69(17), 4215-4226.
- 28.Sulpice R, Flis A, A, Apelt F, Krohn N. (2014) Arabidopsis coordinates the diurnal regulation of carbon allocation and growth across a wide range of photoperiods. doi: 10.1093/mp/sst127. , Molecular Plant 7(1), 137-155.
- 29.J, Jiménez A, Camejo D, M C, Lázaro-Payo A. (2013) Dissecting the integrative antioxidant and redox systems in plant mitochondria. Effect of stress and S-nitrosylation. doi: 10.3389/fpls.2013.00460. Frontiers in plant science. 4-460.
- 30.Das K, Roychoudhury A. (2014) Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. doi: 10.3389/fenvs.2014.00053. Frontiers in. , Environmental Science 2, 53.
- 31.Yazdani M, Mahdieh M. (2012) Salinity induced apoptosis in root meristematic cells of rice. doi:. , 10.7763/IJBBB.2010.V2.66.International Journal of Bioscience, Biochemistry and Bioinformatics 2(1).
- 32.Millar H, N L Taylor. (2014) Subcellular proteomics—where cell biology meetstein chemistry. doi: 10.3389/fpls.2014.00055. Frontiers in plant science. 5, 55.
- 33.Shomali A, Aliniaeifard S. (2020) Overview of Signal Transduction. in Plants Under Salt and Drought Stresses. doi: 10.1007/978-3-030-40277-8_9. Salt and Drought Stress Tolerance in Plants,231 .
- 34.Malakshah Nohzadeh, Habibi Rezaei S, Heidari M, M, Salekdeh Hosseini et al. (2007) Proteomics reveals new salt responsive proteins associated with rice plasma membrane. doi: 10.1271/bbb.70027.Bioscience, biotechnology, and biochemistry. 71(9), 2144-2154.
- 35.N C Turner. (2018) Turgor maintenance by osmotic adjustment: 40 years of progress. doi:doi.org/10.1093/jxb/ery181.Journal of experimental botany. 69(13), 3223-3233.
- 36.J R Acosta-Motos, M F Ortuño, Bernal-Vicente A, Diaz-Vivancos P, Sanchez-Blanco.M.J et al (2017). Plant responses to salt stress: adaptive mechanisms. doi: 10.3390/agronomy7010018.Agronomy. 7(1), 18.
- 37.Sharma A, Shahzad B, Kumar V, S K Kohli, Sidhu G P S.et al (2019). Phytohormones regulate accumulation of osmolytes under abiotic stress. doi: 10.3390/biom9070285.Biomolecules, 9(7). 285.
- 38.Iqbal N, Nazar R, Khan N A eds. (2015) Osmolytes and plants acclimation to changing environment: emerging omics technologies.
- 39.P S Reddy, Jogeswar G, G K Rasineni, Maheswari M, A R Reddy. (2015) Proline over-accumulation alleviates salt stress and protects photosynthetic and antioxidant enzyme activities in transgenic sorghum [Sorghum bicolor (L.) Moench]. doi: 10.1016/j.plaphy.2015.05.014.Plant Physiology and. , Biochemistry 94, 104-113.
- 40.Roychoudhury A, Banerjee A. (2016) Endogenous glycine betaine accumulation mediates abiotic stress tolerance in plants. , Trop Plant Res 3, 105-111.
- 41.Anselment B R K. (2012) Optimization andModelingof Protein Refolding Conditions (Doctoral dissertation,TechnischeUniversitätMünchen).
- 42.I G Mishra, Sharma A. (2012) Exogenously supplied osmoprotectants confer enhanced salinity tolerance in rhizobacteria. , Journal ofEcobiotechnology
- 43.D M Almeida, M, N J Saibo. (2017) Regulation of Na+ and K+ homeostasis in plants: towards improved salt stress tolerance in crop plants. doi: 10.1590/1678-GMB-2016-0106.Genetics and molecular biology. 40(1), 326-345.
- 44.J R Acosta-Motos, M F Ortuño, Bernal-Vicente A, Diaz-Vivancos P, M J Sanchez-Blanco.et al (2017). Plant responses to salt stress: adaptive mechanisms. doi: 10.3390/agronomy7010018. , Agronomy 7(1), 18.
- 45.M R Saric, Loughman B C eds. (2012) Genetic Aspects of Plant Nutrition:. Proceedings of the First International Symposium on Genetic Aspects of Plant Nutrition, Organized by the Serbian Academy of Sciences and Arts, Belgrade,8.Springer Science&Business Media .
- 46.Keisham M, Mukherjee S, S C Bhatla. (2018) Mechanisms of sodium transport in plants—progresses and challenges. doi: 10.3390/ijms19030647. International journal of molecular sciences. 19(3), 647.
- 47.Pang Q, Chen S, Dai S, Chen Y, Wang Y.et al (2010). Comparative proteomics of salt tolerance in Arabidopsis thaliana and Thellungiella halophila. doi: 10.1021/pr100034f.Journal of proteome research. 9(5), 2584-2599.
- 48.Slama I, Abdelly C, Bouchereau A, Flowers T, Savouré A. (2015) Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. doi: 10.1093/aob/mcu239.Annals of Botany. 115(3), 433-447.
- 49.Chen M, Yang Z, Liu J, Zhu T, Wei X et al. (2018) B., Adaptation mechanism of salt excluders under saline conditions and its applications. doi:. 10.3390/ijms19113668. International journal of molecular 19(11), 1-13.
- 50.Shabala S. (2013) Learning from halophytes: physiological basis and strategies to improve abiotic stress tolerance in crops. doi: 10.1093/aob/mct205.Annals of botany. 112(7), 1209-1221.
- 51.D V Assaha, Ueda A, Saneoka H, Al-Yahyai R, M W Yaish. (2017) The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Frontiers in physiology. 8, 509.
- 52.Krishnamurthy A, Rathinasabapathi B. (2013) Oxidative stress tolerance in plants: novel interplay between auxin and reactive oxygen species signaling. doi: 10.3389/fphys.2017.00509. , Plantsignaling&behavior 8(10), 257-261.
- 53.Basu S, Ramegowda V, Kumar A, Pereira A. (2016) . Plant adaptation to drought stress. doi: 10.12688/f1000research.7678.1. F1000Research, 5
- 54.R O Magwanga, Lu P, J N Kirungu, Lu H, Wang X. (2018) Characterization of the late embryogenesis abundant (LEA) proteins family and their role in drought stress tolerance in upland cotton.BMCgenetics,19(6):. 1-31.
- 55.Liu Y, Liang J, Sun L, Yang X, Li D. (2016) . Group3 LEA protein, ZmLEA3, is involved in protection from low temperature stress.Frontiers in plant science,7: 1-10.
- 57.Hanin M, Brini F, Ebel C, Toda Y, Takeda S. (2011) Plant dehydrins and stress tolerance: versatile proteins for complex mechanisms. doi: 10.4161/psb.6.10.17088.Plant signaling &. , behavior 6(10), 1503-1509.
- 58.K M Balmant, Zhang T, Chen S. (2016) Protein phosphorylation and redox modification in stomatal guard cells. doi: 10.3389/fphys.2016.00026. Frontiers in physiology. 7-26.
- 59.Zhao Z, Zhang W, B A Stanley, S M Assmann. (2008) Functional proteomics of Arabidopsis thaliana guard cells uncovers new stomatal signaling pathways. doi: 10.1105/tpc.108.063263.ThePlant Cell,20(12):. 3210-3226.
- 60.M B Bogeat-Triboulot, Brosché M, Renaut J, Jouve L, Le.Thiec, D.,et al (2007). Gradual soil water depletion results in reversible changes of gene expression, protein profiles, ecophysiology, and growth performance inPopuluseuphratica, a poplar growing in arid regions. doi: 10.1104/pp.106.088708.Plantphysiology, 143(2) :. 876-892.
- 61.Joshi R, S H Wani, Singh B, Bohra A, Z A Dar. (2016) Transcription factors and plants response to drought stress: current understanding and future directions. doi: 10.3389/fpls.2016.01029. Frontiers in Plant Science. 7-1029.
- 62.Hou Q, Ufer G, Bartels D. (2016) Lipid signalling in plant responses to abiotic stress. doi: 10.1111/pce.12666.Plant, cell & environment. 39(5), 1029-1048.
- 63.D B Gupta, Rai Y, Gayali S, Chakraborty S, Chakraborty N. (2016) Plant organellar proteomics in response to dehydration: turning protein repertoire into insights. Frontiers in plant science. 7-460.
- 64.Chourey K, Ramani S.and Apte, S.K.(2003).Accumulation of LEA proteins in salt (NaCl) stressed young seedlings of rice (Oryza sativaL.) cultivar Bura Rata and their degradation during recovery from salinity stress. doi: 10.1078/0176-1617-00909.Journal of plant physiology. 160(10), 1165-1174.
- 65.Zang X, Komatsu S. (2007) A proteomics approach for identifying osmotic-stress-related proteins in rice. , Phytochemistry 68(4), 426-437.
- 66.B R Chitteti, Peng Z. (2007) Proteome and phosphoproteome dynamic change during cell dedifferentiation. in Arabidopsis. doi: 10.1016/j.phytochem.2006.11.005. Proteomics 7(9), 1473-1500.
- 67.G H Salekdeh, Siopongco J, L J Wade, Ghareyazie B, Bennett. (2002) analysis of rice leaves during drought stress and recovery. doi: 10.1002/1615-9861 (200209)2: 9<1131::AID-PROT1131>3.0.CO; 2-1.PROTEOMICS:. , International Edition 2(9), 1131-1145.
- 68.Hammami H, Saadatian B, Hosseini S A H. (2020) Geographical variation in seed germination and biochemical response of milk thistle (Silybum marianum) ecotypes exposed to osmotic and salinity stresses.Industrial Crops and Products,152. 112507-10.
- 69.Lin J, Dang F, Chen Y, Guan D, He S. (2019) CaWRKY27 negatively regulates salt and osmotic stress responses in pepper.Plant. , Physiology and Biochemistry 145, 43-51.
- 70.Wang H, Dong Q, Duan D, Zhao S, Li M et al. (2018) Comprehensive genomic analysis of the Tyrosine aminotransferase (TAT) genes in apple (Malus domestica) allows the identification of MdTAT2 conferring tolerance to drought and osmotic stresses in plants.Plant Physiology and Biochemistry,133. 81-91.
- 71.Jin M, Wang H, Liu H, Xia Y, Ruan S et al. (2020) Oxidative stress response and proteomic analysis reveal the mechanisms of toxicity of imidazolium-based ionic liquids against Arabidopsis thaliana.Environmental Pollution. 114013.
- 72.Tanveer M, Shahzad B, Sharma A, Biju S, Bhardwaj R. (2018) 24-Epibrassinolide; an active brassinolide and its role in salt stress tolerance in plants: A review.Plant Physiology and. , Biochemistry 130, 69-79.
- 73.Derouiche A, Cousin C, Mijakovic I. (2012) Protein phosphorylation from the perspective of systems biology. , Curr Opin Biotechnol 23, 585-590.
- 74.Hashiguchi A, Komatsu S. (2016) . Impact of Post-Translational Modifications of Crop Proteins under Abiotic Stress. Proteomes 4(4), 42.
- 76.F Gong Wu, Cao D, Hu X, Wang W. (2016) Advances in crop proteomics: PTMs of proteins under abiotic stress. Proteomics 16(5), 847-65.
- 77.Brader G, Djamei A, Teige M, Palva E T, Hirt H. (2007) The MAP kinase kinase MKK2 affects disease resistance in Arabidopsis. , Mol Plant Microbe Interact 20, 589-596.
- 78.Wang K, Yang Z, Qing D, Ren F, Liu S et al. (2018) Quantitative and functional posttranslational modification proteomics reveals that TREPH1 plays a role in plant touch-delayed bolting. , PNAS 115 43, 10265-10274.
- 79.Hotamisligil G S, R J Davis. (2016) Cell Signaling and Stress Responses. , Cold Spring Harb Perspect Biol. Oct; 8(10), 006072.
- 80.Koeberle A, Pergola C, Shindou H, Koeberle S C, Shimizu T et al. (2015) Role of p38 mitogen-activated protein kinase in linking stearoyl-CoA desaturase-1 activity with endoplasmic reticulum homeostasis. , FASEB J 29(6), 2439-49.
- 81.Katagiri T, Takahashi S, Shinozaki K. (2001) Involvement of a novel Arabidopsis phospholipase D, AtPLD delta, in dehydration-inducible accumulation of phosphatidic acid in stress signaling. , Plant J 26, 595-605.
- 82.J K Zhu. (2002) Salt and Drought Stress Signal Transduction in Plants. , Annu Rev Plant Biol 53, 247-273.
Cited by (4)
This article has been cited by 4 scholarly works according to:
Citing Articles:
Gesunde Pflanzen (2023) Crossref Semantic Scholar OpenAlex
Plant Genetic Researches (2022) OpenAlex
A. Hossain, S. Maitra, B. Pramanick, K. Bhutia, Z. Ahmad et al. - Plant Perspectives to Global Climate Changes (2022) Semantic Scholar OpenAlex
Agronomy (2021) Crossref Semantic Scholar OpenAlex