
Abstract
The effects of long-term space flight on human triceps surae (TS) muscle function and electromechanical delay (EMD) have been investigated. Voluntary and electrically evoked contractions of the TS were obtained from 7 male cosmonauts 30 days before and 3 days after landing. For all cosmonauts the isometric maximal voluntary contraction was reduced by 41.7 % (p < 0.01), whereas the electrically evoked maximal tetanic contraction force (Po) was found to decrease by 25.6 % (p < 0.05). Force deficit increased by 50 % (p < 0.001). This suggests that most of the force loss is due to a reduction in motor drive (motor control). The decrease in Po was associated with a significant increase of the corresponding maximal rates of tension development (43.7 %). The twitch tension (Pt) was not significantly changed and the Pt/Po ratio was increased by 46.7 % (p < 0.05) after space mission. The twitch time-to-peak tension of the TS increased by 7.7 %, but half-relaxation time decreased by 20.6 %. Force-velocity properties of the TS calculated according to a relative scale of voluntary contraction development significantly decreased. The calculations of the same properties of electrically evoked contraction development did not differ substantially from the initial physiological state. Total reaction time (TRT), pre-motor time (PMR) and motor time or EMD were determined. In response to a supramaximal single electrical pulses applied to the tibial nerve, the latent period between the M-wave and Рt beginning was determined. The voluntary contraction EMD increased by 34.1 %; but PMR and TRT decreased by 19.0 and 14.1 %, respectively. The EMD of electrically evoked contraction did not significantly change. Thus, the comparison of the mechanical alterations recorded during voluntary contractions and in contractions evoked by electrical stimulation of the motor nerve, suggests that weightlessness not only modifies the peripheral processes associated with contractions, but also changes central and/or neural command of the contraction.
Author Contributions
Academic Editor: Tetsuro Tamaki, Tokai University School of Medicine
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2017 Yuri A. Koryak
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Competing interests
The authors have declared that no competing interests exist.
Citation:
Introduction
Many properties of the skeletal muscles are known to be altered by an exposure to conditions of weightlessness. Observations from the «Skylab» missions, and more recently the US «Spacelab» and Soviet/Russian Space Station «MIR» missions, have shown weightlessness not only to induce muscle atrophy and decline of muscle strength, but also to reduce the physical work capacity and increase the fatigability of the crew 18. In prolonged space travel, these contractile properties alterations could limit the crew’s ability to work in space and/or on the surface of Mars and to rapidly egress in an emergency on return to Earth. The countermeasures employed during space flights have not been completely successful in preventing reductions and alterations in neuromuscular performance 23, 29, 57.
Changes in contractile properties produced by microgravity are complex. Numerous observations have supported these initial expectations. It was clear from the earliest short-term flights that neuromuscular function was negatively affected by flight 2, 18, 21, 32, 68. After a 17-day «Shuttle» space flight (STS-78), Narici et al. 68 studied muscle function shown that maximal voluntary contraction (MVC) of the plantar flexors was not significantly altered during space flights, the peak tetanic torque elicited by 50 impulses⋅s−1 direct electrical stimulation showed a decline during space flight (10 %). Widrick et al. 86 determined the peak force of individual slow type I and fast type IIa fibers of the soleus and gastrocnemius muscles in four crewmembers on the same space flight (STS-78). The peak force dropped by 21 % in type I fiber and by 25% in type IIa fibers from the soleus muscle. The single fibers from the gastrocnemius (both type I and IIa) were less affected than the soleus muscle fibers 25. The authors suggest that plantar flexor force is lost by 17 days in space, and that the soleus muscle contributes more to the decline than the gastrocnemius. «Skylab-2» (28-day mission) showed a greater drop in thigh vs. arm and extensor vs. flexor torque, with the peak extensor torque of the thigh declining by 20 % compared with a 10% loss in thigh flexor and arm extensor groups. «Skylab-3» (59-day mission) showed an even greater difference between the microgravity-induced loss in thigh (mean 20 % drop) and arm torque (mean 2 % drop) than Skylab-2. The differences were ameliorated on «Skylab-4» (84-day mission), in which the leg exercise countermeasures were credited with reducing the mean loss of peak thigh torque to 6 % 32.
Similar neuromuscular functional tests were performed during long-term space flights 2, 4, 55, 57. In longer «MIR» flights of 6-mo duration, three cosmonauts shown a 20-48 % decline in the MVC of calf plantar flexion 89. After 237-days in space flight, both ankle dorsiflexion and plantar flexion strength were significantly reduced 34. Antonutto et al. 5 demonstrated peak force during two-legged extension to decline by 26 and 29 % after 169, and 180-days in microgravity, respectively. In the previous study 45, 47, 55, 57 we have shown that long-term voluntary bed rest (a 120-day) or long-term space flights (a more 120-days) of the triceps surae muscle produced a significant increase in the time-to-peak tension, and a reduction in MVC, and the maximal strength of a stimulated contraction. The decline contractile properties (in muscular strength) observed in «Skylab» astronauts and Soviet/Russian cosmonauts occurred despite in-flight countermeasures (e.g., treadmill, cycle ergometer, lower body negative pressure). Decrements in motor performance could have negative implications for effective completion of mission-critical operational tasks. None of these studies, however, provide any insight as to whether the space flights-induced decrements in neuromuscular function were of a neural or a muscular origin.
Surface electromyogram (EMG) data show the electrical activity of muscle and have been used in the analysis of human movement. The electromechanical delay (EMD) is traditionally defined as the time lag between the onset of muscle electrical activation and the onset of force production 15. Muscle force is registered only when the contractile elements of the muscle shorten, thus stretching the series elastic components (SEC), which participates in the transmission of force to tendons and joints 38. If the contraction strength changes, significant changes are seen in the EMD values and, therefore, in the SEC 65. This confirms the idea that the time until the SEC stretches is a primary EMD determinant. Furthermore, the SEC structures, originally consisting of an active part (located in the myofibrilare) and a passive one (represented mainly by the aponeurosis and tendon) 88, may contribute to the EMD value.
The EMD changes can be attributed to changes in muscle SEC stiffness. Muscle stiffness is described as the ratio between the force and length of stretching. A mechanically stiff muscle will transmit more force if the SEC stretches a small amount and, on the contrary, mechanically “gentle” (or “slack”) tissue requires a significantly stronger muscle contraction in order to stretch the SEC and produce measurable force. “Slack” tissues require more time from the moment of activation to the start of force generation, i.e. their EMD is longer and it is shortened when muscle stiffness is increased before muscle tension 67. Therefore, the EMD may become a criterion for the differences in stiffness of the musculo-tendinous complex (MTC) under different conditions.
Musculo-tendinous stiffness (MTS) is known to increase after of strength training 35, and it may therefore be expected that the EMD would be shorter in those subjects who practiced physical training in microgravity conditions 54, compared to those who were not subject to physical training 53.
Most of the information on unloaded muscle function in humans has been obtained from ground-based simulation of weightlessness (i.e., bed rest, unilateral lower limb suspension, «dry» water immersion). These studies have shown that muscle strength is reduced by 12-30 % 1, 2, 34, 44, 45, 50. These studies have shown changes in muscle that appear to mimic those observed with 0 G 6. However, studies providing information that directly compares skeletal muscle characteristics during space flights and ground-based analogs of simulated weightlessness have not been conducted. We studied seven crew members before and after of long-term space flights (a more 120 days) Space Station «MIR» mission. We also studied eight subjects after of a 120-days strict bed rest in an antiorthostatic position (-6º) head-down tilt in normal adult subjects 47, 49.
The work presented here is the first study to make quantitative measurements of the contractile properties of the skeletal muscle in men exposed to long-term space travel. The investigation was concerned with the mechanical responses of the triceps surae (TS) muscle. The calf muscle was chosen because it is believed to be one of the skeletal muscles most affected by microgravity unloading because of its postural role in a 1-G environment 13. Mechanical and electrical parameters were recorded during electrical stimulation of the muscle motor nerve to distinguish peripheral changes from those occurring centrally.
Up to now, due to methodological difficulties, examination of the intrinsic contractile properties of human skeletal muscles in a true weightless environment and/or during its simulation has been beyond the capability of scientists who have therefore concentrated mostly on investigating the mechanical features of the voluntary muscle contractions. The primary goal of the present work was to use single muscle to determine whether mechanisms of human muscle contraction were affected by prolonged space flight. Since we had previously used identical methodology to investigate the effects of a long-term period of bed-rest on the human TS muscle function 36, 42, our secondary goal was to test the validity of ground-based bed-rest as a model of human space flight.
Methods
The experimental protocol was approved by the Russian National Committee on Bioethics of the Russian Academy of Sciences and the Yu. A. Gagarin Cosmonauts Training Center at Start City and was in compliance with the principles set forth in the Declaration of Helsinki (last modified in 2004).
Participants
The subjects in this study were seven male cosmonauts who participated in Space Station «MIR» missions («MIR» missions EP 18, 22 24 2627). Prior to the flight, their average age, height, body mass were 45.1 ± 2.0 years (range 43-54), 176.0 ± 2.3 cm (range 167-182), 79.9 ± 2.0 kg (range 70-86), respectively (Table 1). The duration of the space flight was by mean 213.0 ± 30.5 days (range 115-380). Each subject performed two sets of experiments 30 days before space flight (baseline data collections) and immediately after space flight (return) of 3 days.
Table 1. Physical characteristics (М m, n = 7) of the permanent crewmembers of space mission| Age, (years) | Height, (cm) | Body mass, (kg) | Duration space, (day/hour/min) |
| 54 | 167 | 79.3 | 115/08/43 |
| 40 | 182 | 83 | 196/16/31 |
| 50 | 174 | 86 | 379/14/51 |
| 43 | 182 | 81 | 196/17/26 |
| 40 | 175 | 83 | 196/17/26 |
| 47 | 170 | 77 | 0207/12/02 |
| 42 | 182 | 70 | 188/20/16 |
| 45.1 ± 2.0 | 176.0 ± 2.3 | 79.9 ± 2.0 | 213.0 ± 30.5 |
Familiarization
The cosmonauts reported to the laboratory twice. During the first visit, the cosmonauts were familiarized with the experimental set-up and procedures. The cosmonauts gave their written informed consent to participate in this study. On a subsequent day, the cosmonauts were involved in testing contraction properties of the ankle extensor muscles. Each subject served as his own control.
Testing Procedure and Measurement
Isometric Set-up
The mechanical responses of the TS muscle were recorded by method tendometry (59) (Figure 1, top panel, left) which made it possible to measure the force of a single muscle contraction by the degree of tension change in muscle distal tendon (44, 46-48). Measurement of muscle tension using a strain-gauge transducer is based on the physical law of the resolution of forces according to the parallelogram principle (Figure 1, top panel, insert). If a strain-gauge transducer is pressed to the tendon, the transducer causes it to bend at an angle. The force (F1) that is directed along the muscle axis to the proximal point of attaching and originates during the muscle contraction is oppositely directed and equal to the force (F2) that is directed to the distal point of the tendon attachment. F1 which is directed across the tendon, operates at the point of the transducer and tendon contraction. If the angle at which the tendon bends is constant, the force (F) recorded by the strain-gauge dynamometer is proportional to F1 (or F2). A rigid dynamometer is needed for recording the muscle force using a strain-gauge transducer, because any deformation under tendon pressure will change the transducer position and alter the tendon angle. A steel dynamometer ring was used in our transducer.
The subject was seated comfortably on a special chair in a standard position (knee joint angle between tibia and sole of foot of ~ 90 deg). The position of the seat was adjusted to the individual and then firmly secured. A rigid leg fixation provided the isometric regimen of the muscle contraction. The dynamometer that is a steel ring with a saddle-shaped special block was tightly attached to support the Achilles tendon of the muscle. The degree of pressure between the tendometrical sensor and the tendon was constant for all the subjects and amounted to 5 kg.
Surface Electromyography
After careful preparation of the skin including shaving excess hair, abrading the skin with fine sandpaper, and cleaning the skin with an isopropyl alcohol swab to reduce impedance below 5 kΩ, pairs of surface Ag-AgCl electrodes (∅ 8 mm, center to center distance 25 mm and a recording area of 50 mm2), were placed over the muscle bellies of medial (MG) and lateral (LG) gastrocnemius, and soleus (Sol) muscles. The electrodes were placed longitudinally with respect to the underlying muscle fiber arrangement and located according to the recommendations by SENIAM (surface EMG for noninvasive assessment of muscles). For the soleus muscle, electrodes were attached to the skin over the middle part of the soleus muscle, belly and placed ~ 6 cm distally of the stimulation electrodes The large grounding electrode (7.5 cm x 6.5 cm) was located on the proximal portion of the leg between the upper recording electrode and the stimulating electrodes. Recordings of the electrical responses (EMG) of the skeletal muscle during slow MVC were made during for a short period (~ 0.2 s). EMG signals were amplified (x 1,000) and digitized (bandwidth of 0–2,000 Hz) at a sampling rate of 5 kΩ. During contraction the EMG signal was recorded on FM tape and simultaneously displayed on an oscilloscope to assess its quality. EMG signals were bandpass filtered between 20 and 500 Hz, and smoothed using a moving root-mean- square filter with a time constant of 50 ms.
Electrical Stimulation
A single, square-wave, supramaximal transcutaneous electrical stimulus of 1 ms duration was delivered to the n. tibial. Transcutaneous electrical stimuli were delivered to the tibial nerve using a high-voltage (maximal voltage = 400 V), constant current stimulator (model «ESU-1», USSR). The cathode (active electrode) was a metal probe (∅ 1 cm) with the tip covered in a saline-soaked sponge, which was pressed over the tibial nerve in the popliteal fossa which is the place of the lowest resistance. The anode (passive electrode) was a 6 x 4 cm rectangular self-adhesive electrode that was positioned between the tibial tuberosity and the patella. The large grounding electrode (7.5 cm x 6.5 cm) was located in the proximal portion of the leg between the peak-up and stimulating electrodes. Single stimuli were administered to the tibial nerve at a low current (amperage = 20 mA) to determine the optimal probe location based on the visual inspection of the compound muscle action potential (M-wave) and H-reflex of the soleus muscle that were monitored on an cathode-ray storage tube oscilloscope. A single stimulus was given every 30 s. Once the location was determined and marked, the maximal M-wave was achieved with incremental (5 mA) amperage increases until a plateau in the peak-to-peak M-wave was observed after three successive amperage increases. To assure a supramaximal stimulus, 120 % of the stimulus that elicited the maximal M-wave was used during the evoked twitch procedures.
The isometric tetanic contractions of the TS muscle were induced by electrical stimulation of the n. tibial using supramaximal rectangular pulses with a frequency of 150 impulses⋅s−1 (52). All the recordings were made in a room at constant temperature (~ 22 ± 1 C).
The contractile properties of the TS muscle were tested twice: pre- (~ 30-days) and post-space flight (~ 3-4-days) and the test protocol were identical for both pre mission and post mission tests.
Tension Properties
The whole protocol was executed by one investigator. For all cosmonauts, the right leg was studied. The contractile properties of the TS muscle were estimated according to the mechanical parameters of a voluntary and electrically evoked contraction (maximal isometric twitch and tetanic contractions, double stimulation).
Voluntary Contraction
The maximal voluntary contraction was measured from the tendogram of an isometric voluntary contraction performed, on instruction, «to exert a maximal contraction» (Figure 1, top panel, right). Maximal voluntary contraction was determined during three contractions of 3-4 s duration separated by 3 min. The largest of three contractions was considered as the maximal voluntary contraction (MVC). During the contractions, the subjects were verbally encouraged and a visual feedback was provided.
Evoked (tetanic) Contraction
The maximal strength was measured from the tendogram from the evoked contraction (Po ,Figure 1, top panel, right).) in response to an electric tetanic stimulation of the nerve, innervating the TS muscle, with a frequency of 150 impulses⋅s−1 (43, 44).
The difference between Po and MVC expressed as a percentage of the Po value and referred to as force deficit (Pd) has also been calculated (43, 44) (Figure 1, top panel, right).
Evoked (double) Contraction
Duration of an active state plateau of the contractile components of muscle fibers of the TS muscle was measured using the method of interval between stimuli. Supramaximal twin stimuli at 330, 250, 200, 100, 50, and 20 Hz were applied (46). The maximal strength (amplitude) of the muscle contraction was determined and expressed as a percentage of the twitch contraction.
Velocity Properties
The maximal isometric peak twitch force (Pt) was measured from the tendogram of the ts muscle isometric twitch response to a single electrical stimulus applied to the tibial nerve. The time from the moment of stimulation to peak twitch (TPT), the time from contraction peak to half-relaxation (1/2 RT) and total contraction time (TCT) - the time from moment of stimulation to the total muscle relaxation - were calculated from the tendogram of the isometric twitch (Figure 1, top panel, right).
Rate of Force Development
The rate of development of increased muscle tension was calculated from the tendogram by the isometric voluntary contraction after the instruction «to exert the fastest and greatest tension» using a relative scale, i.e. the time of reaching 25, 50, 75, 90 % of maximum tension (36, 37, 43, 44). Similarly, the rate of rise of the evoked contraction, in response to electrical stimulation of the n. tibial with a frequency of 150 impulses⋅s−1 (43, 44) (Figure 1, top panel, right). The maximum rates of voluntary contraction (dPvc/dt), and evoked contraction (dPec/dt) development were obtained by differentiation of the analog signal.
Electromechanical Delay
Electromechanical delay (EMD) comprises an important portion of the rate of tension development phase of the contraction (28), where changes in motor unit activity patterns have been demonstrated to play a critical role in the maximal rate of force development (84).
The experiments were divided into two protocols.
Protocol 1 ⎯ Voluntary Contraction
On a light signal the subject carried out plantar flexor under condition of «to contract as it is possible quickly and strongly» (Figure 1, bottom panel). Voluntary contraction in response to a visual stimulus (flash lamp) was adopted as a rapid ballistic movement. The signal to movement of «explosive» character was the visual diode − lamp (Ø 7 mm, 1 W) ⎯ was placed at eye level 1 m in front of the subject. Lasted signals were 2.5 s and the pause between the signals was random ranging from 1.4 to 5.0 s. The threshold for force was 5 N.
A separate timer was used to record the time interval from the presentation of the light signal to movement. The special timer allowing synchronously with presentation of a light signal to the beginning of movement to carry out record of development of mechanical answer the human TS muscle was used.
From the tendogram estimated total reaction time (TRT), defined as the time interval from the application of the light stimulus to movement. TPT was divided into pre-motor (PMT), defined as the time interval from the application of the stimulus to the change in electrical activity of the soleus muscle. EMD or motor time (MT) was determined from the time lag between the onset of dorsiflexion force and surface EMG in the soleus muscle (84). Surface EMG onset threshold was identified as the first point of the surface EMG signal to rise above the 95 % confidence interval for baseline noise and to remain above the 95 % confidence interval for 20 ms (22). The force thresholds were also taken as relative values of 2 % from the maximum isometric force level of each contraction. The onset of EMG integration was initiated 70 ms before the identified onset of the force time curve, which accounts for EMD (8).
Subjects were permitted three practice trials separated by 30 s and in most cases the mean of three readings was used to determine TRT, PMT and EMD.
Protocol 2 ⎯ Twitch Contraction
To evaluate time of involuntary EMD, rectangular electrical pulses were applied to n. tibialis. The active electrode was placed over the popliteal space. Four single supramaximal stimuli were delivered to n. tibialis and a maximal M-wave recorded. The rest interval between stimuli was no less than 30 s. The EMD was determined as a time interval between stimulation artifact and the onset of the twitch response (Figure 1, top panel, right, insert).
Figure 1.Experimental set-up and scheme estimated of parameters. A. Experimental set-up and scheme the principle of tendometry method: B. Examples measurements of mechanical response parameters in isometric twitch contraction curve (left) and in electrically evoked tetanic tension and voluntary muscle tension development (right), and measurements electromechanical delay (insert) TPT, a time-to-peak; 1/2RT, a half-relaxation time; TCT, a total contraction time; Pt, a twitch force; EMD, a electromechanical delay. C. Schematic presentation of a sample contraction showing total reaction time (TRT) with its premotor (PMT) and motor (MT) components, force-time curve and EMG recorded from m. soleus.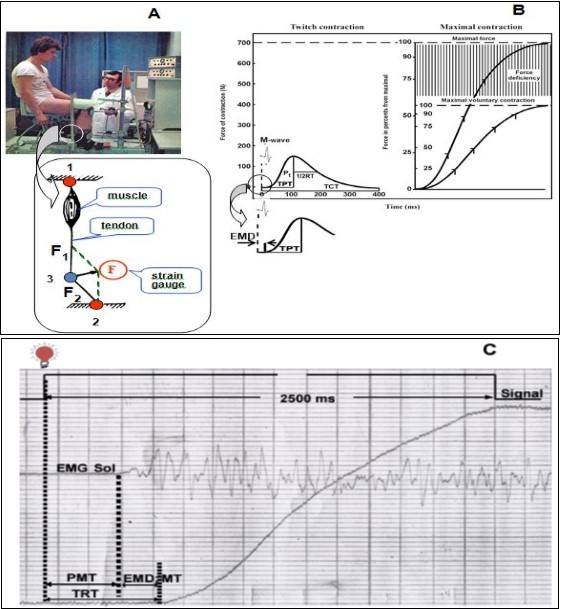
The results of the experiment were simultaneously recorded on magnetic tape and simultaneously displayed on an oscilloscope to assess its quality.
Exercise Regimens
To preserve the muscle contractile properties in long-term space missions, cosmonauts used physical training (PT). Details of the training program and performance tests have been provided elsewhere 24, 76. PT which were mostly of a locomotor type and included a warm-up (walking on a treadmill for 5 min), and low (2 min), moderate (2 min) and maximum (1 min) intensity running. PT were performed for 1.5–2 hours. Four-day microcycles, each including 3 days with PT and 1 day of rest, were followed in the PT program 24, 76.
A treadmill and a cycle ergometer were the main PT tools, training the cardiovascular and respiratory systems 33. The treadmill device could be used in a passive (subject driven) or active (motorized) mode of operation, which was selected by the crewmember during each exercise session. Crewmembers used a subject-loading device to fix themselves to the treadmill, which provided varying levels of loading relative to body weight (typical load was ~ 70 % of body weight) during use. In this way, the crewmembers could complete running or walking exercise while partially loaded. The cycle ergometer provided a load of 50–225 W at a pedaling rate of 40–80 rpm. The crewmembers also had access to bungee cords, which they could use to provide resistance-type exercise for various muscle groups.
Statistics
Conventional statistical methods were used for the calculation of means and standard errors (± SE). A Wilcoxon's signed-rank test for paired differences was used to test changes in all mechanical parameters due to space flight. Furthermore, differences between baseline (background) values of the subject and those post-space flight were tested for significance by Student′s paired t test. All data are expressed as means ± standard error (SE) within the text and the tables and as means ± SE in the figures for the sake of clarity. The percentage changes for pre- and post-space flight was also calculated.
For comparative change of contractile properties the TS muscle during long-term space flight and observable changes during long-term models simulating weightlessness − bed rest (Koryak 1995a; 1998) similar duration used calculated by ((before ⎯ after) / before) x 100 %).
All statistical analyses were conducted at a level of significance of p < 0.05.
Results
Tension Properties
Mean data of the changes in the TS muscle tension properties are shown in Figure 2 (top panel). Isometric Pt increased by mean 14.8 % (pre 106.0 ± 7.8 N compared to post 121.6 ± 17.6 N) after space flight (p > 0.05). MVC decreased by a mean of 41.7 % (pre 520.0 ± 66.7 N compared to post 303.1 ± 50.0 N; p < 0.05), and isometric muscle tetanic (Po) decreased by a mean of 25.6 % (pre 738.6 ± 65.7 N compared to post-space flights 549.4 ± 37.3 N; p < 0.05), after space flight, respectively. The Pd (Figure 2, top panel) increased significantly by a mean of 49.7 % (pre 32.2 ± 4.6 % compared to post-space flight 46.7 ± 5.4 %; p < 0.01).
Figure 2.The effects of long-term space flights on strength properties A. The maximal twitch response of force (Pt), maximal voluntary contraction (MVC), maximal electrically evoked tetanic contraction (Po, recorded at 150 impulses.s-1) (left panel), and force deficit (Pd,) before and after prolonged space flights (right panel). B. The histograms represent the individual MVC (top panel) and Po (bottom panel) values of each cosmonaut from baseline data collections and 3 days after return test. All values are means ± SE. Significantly different between groups for corresponding contractions: **p < 0.05 ***p < 0.01
A summary of individual values is given in Figure 2 (middle panel). Post-flight maximal isometric MVC in plantar flexion, collected near landing (immediately after space flight (return - R); R+3 days), indicated a decrease in MVC. More precisely, decreases in MVC were found for each cosmonaut from baseline data collections ranging from 22 % to 65 %, respectively). For the population MVC was found to be significantly higher in pre-flight condition than in post-flight condition (p < 0.05).
Post-flight maximal isometric muscle tetanic (Po) in plantar flexion indicated a decrease in Po (Figure 2, lower panel). More precisely, decreases in Po were found for each cosmonaut from baseline data collections (baseline data collections) ranging from 8 to 41 %, whereas 2 subjects (1, and 7, respectively) had had concerning smaller decrease Po (8, and 11 %, respectively). For the population Po was found to be significantly higher in pre-flight condition than in post-flight condition.
The analysis of the data shows, that there is no connection between basic value of force of reduction of a muscle and degree of drop force properties muscle after space flight. For example, cosmonauts 3 and 4, the highest parameters, having up to space flight, MVC, after space flight differ on depth of drop of force of reduction. There are no such connections and for a parameter Po (Figure 2).
Figure 3 shows a 55 % increase (p < 0.05) of iEMG and a 71 % increase (p < 0.05) of the iEMG/MVC ratio for a slow MVC after the space flight. The increase in both IEMG and iEMG/MVC are indicators of decreases in efficiency of the degree of electrical activation in the muscle after flight. The degree of activation required to generate a force is estimated by the EMG/force relation. The slope of the EMG/force relation was decrease significantly after space flight (p < 0.05).
Figure 3. The effects of long-term space flights on EMG activities. Activation of the medial and lateral gastrocnemius and soleus muscles during MVC after long-term space flight. Values are expressed as percentage of control (before). All values are means ± SE. Significantly different between groups for corresponding contractions: **p < 0.05.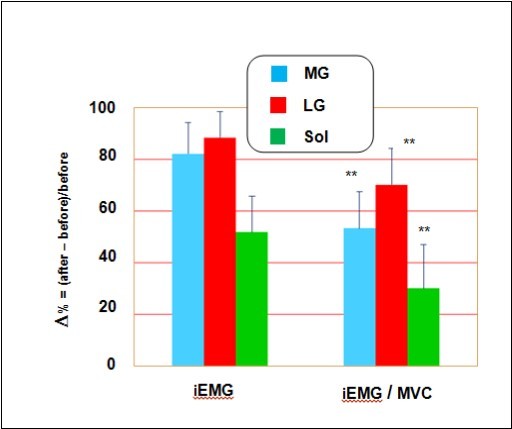
Figure 4.The effects of long-term space flights on maximal electrically evoked contraction with double stimulation. Mean value of the maximal force of contraction of the triceps surae muscle with doublet stimulation at different intervals between impulses. All values are means ± SE. Significantly different between groups for corresponding contractions: **p < 0.05 ***p < 0.01.
The mean changes in force of the ts muscle contraction during doublet stimulation, in which a second pulse was applied at various intervals, are presented graphically in Figure 3. The greatest force of contraction under these conditions was at an interval of between 4 and 10 ms and decrease or increase in the interval were accompanied by a considerable decline. The relative increase in force as a result of doublet stimulation was significantly lower after long-term space flight in comparison with the control (baseline data collections) value (p < 0.01-0.001).
Velocity Properties
The data of the change of mean time of isometric twitch contraction as the opposite value to contraction velocity for the TS muscle after long-term space flight, is given in Figure 5. As is seen from the data analysis, exposure to microgravity conditions was accompanied by a statistically significant decrease of the muscle contraction and increased relaxation (1/2 RT) velocity. Thus, TPT increase by a mean of 7.7 % (pre 130 ± 2 ms compared to post 140 ± 6 ms; p < 0.05) after space flight. 1/2 RT was shorter after space flight than in the control value. Thus, 1/2 RT reduced by a mean of 20.6 % (pre 97 ± 4 ms compared to post 77 ± 4 ms; p < 0.05), but TCT increase by a mean of 7.5 % (pre 456 ± 25 ms compared to post 490 ± 31 ms; p < 0.01).
The same subjects showing increase in TPT also showed significant increase in the twitch-to-tetanic ratio by a mean of 46.7 % (pre 0.15 ± 0.01 compared to post 0.22 ± 0.02; p < 0.05) (Table 2).
Table 2. The mechanical and time characteristics of the human triceps surae muscle after a long-duration space flight (М m, n = 7)| Before | After | ∆ , % | |
| MVCslow, N | 519.9 66.7 | 303.1 50.0 | 41.7 |
| MVCfast, N | 467.9 40.2 | 403.2 43.2 | 13.8 |
| 0,5 MVCfast/t0.5, N/ms | 1.422 1,373 | 1.756 1,67 | 23.4 |
| dPvc/dt, %/ms | 20.60 0 | 13.14 0 | 36.2 |
| Twitch-to-tetanic ratio | 0.15 0.01 | 0.22 0.02 | 46.7 |
| PMT, ms | 256 24 | 208 20 | 18.8 |
| EMD, ms | 31.4 2.8 | 42.1 3.4 | 34.1 |
| Рt, N | 105.9 7.8 | 12.4 1.8 | 14.8 |
| dPt/dt, %/ms | 13.1 0.29 | 1.19 0.02 | 11.2 |
| EMDt, ms | 10.3 0.4 | 12.4 0.3 | 20.4 |
| Ро, N | 738.7 65.7 | 549.4 37.3 | 14.8 |
| dPec/dt, %/ms | 16.38 0 | 23.54 0 | 43.7 |
Individual data of each cosmonaut from baseline data collections and collected near landing (R +3) are summarized in Figure 5 (middle and low panel). Post-flight TPT, collected near landing, indicated a increased in TPT. More precisely, increases in TPT were found for 5 subjects ranging from 8 to 24 %, whereas 2 subjects (3, and 4, respectively) had slightly decreased TPT (6, and 12 %, respectively).
Figure 5.The effects of long-term space flights on velocity properties. Time-to-peak twitch (TPT), and time to half-relaxation (1/2 RT), and total contraction time (TCT) before and after prolonged space flights (top panel), and the histograms represent the individual TPT (middle panel), and made an interval 1/2RT (bottom panel) values of each cosmonaut from baseline data collections and 3 days after return test. All values are means ± SE. Significantly different between groups for corresponding contractions: **p < 0.05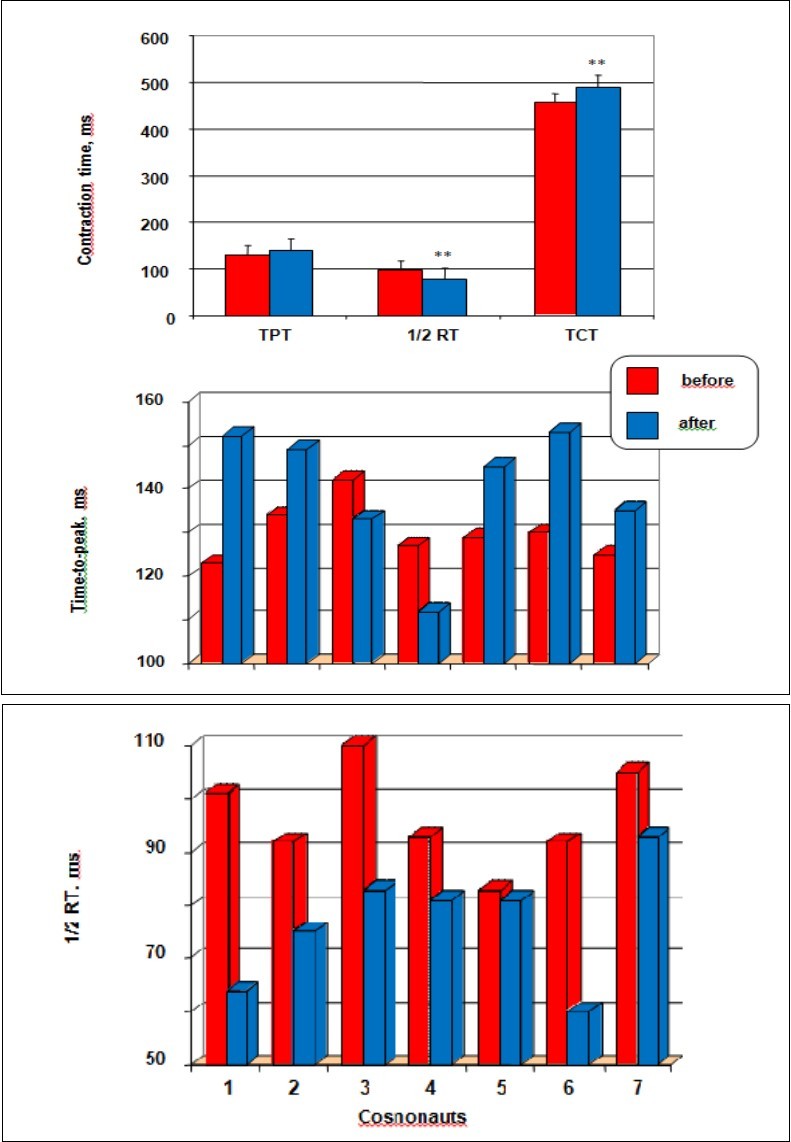
On the other hand, the effect of exposure to microgravity on relaxation (1/2 RT) velocity was tested by comparing 1/2 RT values in pre-flight and post-flight conditions. Seven cosmonauts presented an increase in 1/2 RT ranging made an interval 2 % to 37 %, whereas the one subjects (cosmonaut 5) had slightly decreased 1/2 RT (2 %). Statistical analysis, taking each subject into account, revealed that the decrease in 1/2 RT was significant for the whole population. Individual data are summarized in Figure 5.
Rate of Force Development
Mean changes in the rate of development of isometric tension of the TS muscle are given in Figure 6. The decrease in the MVC (42 %) was associated with a significant slowing of the rate of tension development during a voluntary isometric contraction (Figure 6, left curve; p < 0.01-0.001); maximal dPvc/dt significant decrease by a mean of 36.2 % when normalized as the percentage MVC (Figure 6, insert; p < 0.001).
Figure 6. The effects of long-term space flights on development of force of the triceps surae muscle expressed relative to the maximal force (left) and the maximal rate of rise of tension development (right). Average curves showing the development of force while executing explosive voluntary contraction and as a result of electrical stimulation at 150 impulses×s-1. Significantly different between groups for corresponding contractions: **p < 0.05 ***p < 0.01
Analysis of the force-time curve of the electrically evoked contractions did not reveal significant differences (Figure 6, right curve; p > 0.05) while the maximal dPec/dt was significant increased by a mean of 43.7 % after space flights (Figure 6, insert; p < 0.001). These mechanical dissociations suggested that some state beyond the membrane processes was changed.
Electromechanical Delay
The mean changes in EMD under space flight are shown in Table 2. The EMD increased significantly, by 34.1 ± 1.3 % after space flight (pre 31.4 ± 2.8 ms compared to post 42.1 ± 3.4 ms; р < 0.05).
The PMT decreased significantly by a mean of 18.8 % (pre 256.4 ± 24.1 ms compared to post 207.6 ± 19.6 ms; p < 0.01), and TRT decreased by a mean of 14.1 % (pre 289.1 ± 25.0 ms compared to post 248.4 ± 20.5 ms; p < 0.01) after space flight.
The coefficients of correlation between the EMD and MVC before space flight were −0.67 (p < 0.05) and strongest correlations were found between EMD and ballistic voluntary force (r = 0.90; p < 0.01). The coefficients of correlation decreased between the EMD and MVC after space flight (r = −0.57; p = 0.05), and between the EMD and ballistic voluntary force (r = −0.70; p = 0.07).
EMD during evoked contraction did not differ significantly before and after space flight (Table 2).
Comparison of EMD during evoked and voluntary contractions revealed a longer EMD during voluntary both pre- and post-space flight. For instance, pre-space flight value EMD during voluntary contractions were by a mean of 31.4 ± 3.1 ms in contrast to 10.3 ± 0.4 ms during evoked contractions (p < 0.05); post-flight values were by a mean of 42.1 ± 3.1 ms and 12.43 ± 0.3 ms, respectively (Table 2).
Comparison of Space Flight to a Period of Bed-Rest
In previous work, we examined the responses of the human TS muscle to 120 days of complete bed-rest 44. Figure 7 compares the results from this previous work to the present study. Inter-subject variability appears to be similar for each model. These data indicated no differences in the relative changes between the two models for MVC, and Pt, and Po, and force deficit, TPT and 1/2 RT. On the other hand, our data demonstrate that the countermeasures employed during space flights have not been completely successful in preventing reductions in alterations in neuromuscular performance. A better understanding of how humans respond to space flight is a requirement for the development of more effective countermeasures.
Figure 7.Effects of 120 days of space flight and ground-based bed rest on the functional properties of the triceps surae muscle. The bars represent the mean relative change experienced by cosmonaut and bed rest subjects. Bed rest data obtained from Koryak (44).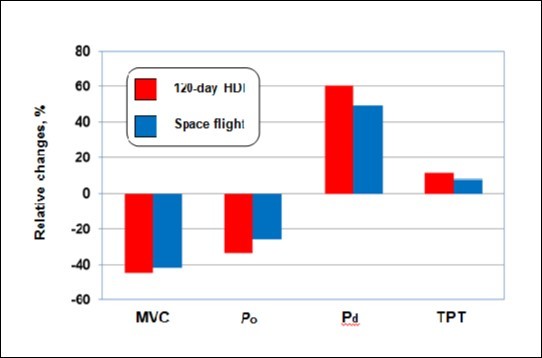
Discussion
The major aim of the present study was to examine the influence of prolonged microgravity (weightlessness) on the contractile responses of skeletal muscle in intact healthy men. Muscle to experimental conditions may occur for a number of reasons and whether the changes in contractile processes or whether the neural (motor) command 43, 73. The results of the study revealed the change in the TS muscle contractile properties after long-term exposure to space flight. The adaptation of muscle contraction to the effect of long-term weightlessness may be the result of changes which occur within the nervous command of muscle contraction or at peripheral sites.
The use of electrically evoked twitch and maximal tetanic contractions is well established in the study of human contractile function. The results of this study demonstrate that contractile properties of skeletal muscle are altered during typical long-term space flights (a more than 4 months) on aboard the Space Station «MIR». Experiments utilizing fixed end, unloading, isometric contractions revealed alterations in force production, velocity and force-velocity output of post-flight muscle. Presumably these alterations result from the lack of normal weight-bearing activity by the lower limb postural muscles in reduced gravity conditions aboard the Space Station.
Effects of Space Flight on Force Properties
We anticipated that the changes in isometric Pt would correspond to those in MVC, namely a decrease with microgravity. In contrast, we observed a nonsignificant increase in Pt after long-term space flights. These unexpected findings are not unique. Edgerton et al. 20 found a nonsignificant increase in Pt after 6 mo hindlimb immobilized skeletal muscles of a nonhuman primate (Galago senegalensis), and Koryak 45, 46 after 7 days ‘dry’ water immersion in the human TS muscle. These changes are quite similar to these observed in our study. These changes in twitch tension may reflect alterations in stiffness (extensibility) in human muscles. A decrease in muscle extensibility (stiffness), which has been observed after disuse/space flight 30, 62 would be the changes in isometric Pt. In contrast, Po decreased after space flight. Thus there is a tendency for the twitch-to-tetanus ratio to increase after space flight. These changes in the twitch-to-tetanus ratio may reflect alterations in muscle extensibility. We speculate that in our experiment, space flight is the cause of a decrease in muscle extensibility.
After prolonged space flights the electrically evoked contractions (Po) decreased significantly at a rate of 26 % of normal. The Po is a direct measure of the force-generating capacity of a muscle and has been considered to reflect the number of active interactions between actin and myosin 17. Disuse produced a decline in Po21, 45, 46. This decline could reflect a decrease in the number of active cross-bridges and be expected to decrease the work capacity. Two hypothesis may be suggested to account for the observation. First, the total number of cross-bridges could have been smaller after the period of space flight. Second, the force output per cross-bridge could have been decreased. However, according to Steven et al. 77 have shown that when it was expressed per cross-sectional area, the force was unchanged after disuse. This would indicate that the first hypothesis of a decrease in the maximal number of cross-bridges was more appropriate to our results, rather than a change in density.
As a whole the mechanisms responsible for the force loss with microgravity are not well understood. However, changes in the force might have been caused by changes in different architectural properties, such as muscle length, and fascicle length, and pennation angles 27, 85. Consequently, the relationships among joint angles, muscle lengths, and pennation angles are highly specific to muscle. Muscle architecture, together with intrinsic properties such as also fiber composition, also affects functional characteristics of muscle, e.g., maximal shortening velocity and Po70. In addition, considering that of the TS muscle our experimental environment was not extended and tensed, which makes possible the associated of such a condition with physiological shortening, it may be assumed that the total number of successively connected sarcomeres was considerably decreased 25. These may have contributed to the reduced in muscle layer thickness and fascicle angles. In fact, according to Clément et al. 16 it seems that cosmonauts adopt a dorsiflexion position during space flights. Then, in postflight, a shortening of the plantar flexors will occur when the preflight neutral position is reached.
The measurement of MVC depends on many factors, such as motivation of the cosmonauts to give their maximal effort. MVC values obtained in pre-flight conditions correspond to the data report in the literature 4, 57, 89. As found by other teams who also worked on the Space Station «MIR» mission 5, 89, our post-flight data indicated for each subject a significant decrease in MVC. For instance, Zange et al. 89, by means of a NMR technique, reported a reduction in muscle mass of the lower limbs of ~ 11 % sand attributed these morphological data to a decrease in muscle cross-sectional area. Our post-flight data indicated for each subject a significant decrease in MVC. The larger fall in MVC in the present study could also be due to the effects of weightlessness on i) the intrinsic characteristics of the recruited motor units and ii) changes in the motor unit recruitment pattern. Such alterations have already been hypothesized by some authors. Additionally, a decline in strength was also proved by Lambertz et al. 62 after a space flight on root mean square values, used for expressing the power of an EMG signal. These studies indicate decrease in normalized mean square values after space flights support this proposition that microgravity interferes substantially with the normal neural drive.
The much larger reduction in MVC (mean decrease 42 %) when compared with to changes in Po after a prolonged space flights of 3 mo on Space Station «MIR» (mean decrease 24 %) may indicate an inability of the central nervous system to activate the TS muscle normally. Whether this was due to a lack of motivation on the part of the subjects, or to an involuntary reduction in neural drive, is difficult to distinguish. Additionally, a decline in maximal power was also proved by Antonutto et al. 5 after 180 days space flight and was attributed to a decrease in neural input. The subjects certainly appeared well motivated and had no discomfort or knee stiffness before performing the test which could have account for the low MVC. The increase in force deficit would suggests a decline in central drive in the control of voluntary muscle by the motor nervous system. In fact, during MVC, the EMG activity has been found to be significantly changed by the inactivity itself 21, 48, 73. Moreover, observation of amplitude changes after inactivity has suggested that fewer motor unit were activated in disused muscle 73, and maximal firing frequency of motor unit has been found to be decreased 21. In fact, this decrease in maximal firing rate should imply the recruitment of a larger number of motor units to develop the same target strength in pre-flight and post-flight conditions.
On the other hand, a decrease in maximal firing rate could be explained by changes in proprioceptive afferents on the motoneurons 64. This suggests that in future studies in human cognizance must be taken of the initial physiological states of the muscle that are to be disused to access the extent to which neural and muscle function is affected by loss of voluntary movement. Additionally, the data of Antonutto et al. 4, 5 suggest that microgravity causes a fundamental alteration in motor control. They observed two-legged muscle power to decline considerably more than could be explained by the loss in muscle mass. Moreover, the loss of explosive leg power was associated with a substantial reduction in the EMG activity of the muscle. The data of Recktenwald et al. 71 also suggest that microgravity induced a reorganization of motor recruitment motor units. These changes of motor control, may be one contributing factor of microgravity to the reduced extensor muscle torque and maximal power previously experienced by cosmonauts/astronauts after space flight 5, 34 and be change in architecture as muscle fibers, and in a muscle-tendon complex, as previously is shown on astronauts after space flight 53 and after stay in head-down tilt bed-rest 41, 60.
During bed-rest, subjects underwent the exact same physiological testing performed by the cosmonauts during this flight. Mechanical responses the TS muscle were obtained and analysed using identical procedures. By directly comparing the results from these two studies we have attempted to assess the validity of the ground-based, head-down tilt bed rest procedure as a model of human space flight. The reductions in both MVC and Po did not vary by more than a few per cent between space flight and bed rest model. Thus, the results of this comparison indicate that in general, mechanisms of force production in TS muscle respond in a similar manner to either bed rest or space flight. We should point out that one difference between the bed rest and space flight studies was that the cosmonauts were required by «MIR» flight surgeons to perform aerobic countermeasure exercise during the flight. It is impossible to evaluate how this may have affected the comparison between bed rest and space flight. We would concur with that conclusion based on the similarities between the bed rest where subjects did not perform exercise countermeasures and space flight results. We studied contractile properties the TS muscle obtained from adult subjects before and after 120 days head down tilt bed rest 44, 47, 49. During bed rest subjects did perform exercise countermeasures. Training had caused a decrease of 3 % in MVC, and Pt, and in Po of 14 %, and of 9 %, respectively. The force deficit had decreased significantly by 10 %. These experimental findings indicated that neural as well as muscle adaptation occurred in response to head-down tilt with countermeasures.
Effects of Space Flight on Velocity Properties
The results showed that a slow contracting muscle group 43, 82 was affected by microgravity. These data extend previous finding concerning the effects of short-term voluntary ‘dry’ water immersion 45, and long-term bed-rest 44, 47 on the mechanical characteristics of the TS muscle. The functional components of the twitch duration are the TPT and the 1/2 RT. Significant increases in maximal isometric twitch TPT and decreased 1/2 RT were maintained during a 120-day bed-rest. In the present study, the difference between the post-flight and pre-flight TS muscles was ~ 8 %. The underlying mechanistic basis of TPT and 1/2 RT involves the competitive interaction of factors involved with activation (Ca2+ release kinetics), cross-bridge cycling, Ca2+ uptake by the sarcoplasmic reticulum (SR), and changes the active fraction (muscle fibers) or the passive fraction (tendons) of the series-elastic component.
A likely explanation of the change in TPT in the real microgravity limbs is a relatively greater atrophy of type I fibres, which has been found to make up the majority of the TS muscle 40. However, since microgravity produces muscle atrophy both in fast and slow skeletal muscles and, in addition, have been shown to cause fibre type-specific changes in the contractile properties 29, other factor(s) may be affected that alter fibre type composition. Additionally, it is tempting to suggest that the decreased 1/2 RT of the space flight TS muscles is probably due to an increased number of fast SR Ca2+-ATP-ase pumps, as observed in previous hindlimb-suspension studies 77.
One additional effect to consider is the fibre type distribution. Is shown, that after space flight percentage of fibers no significantly changed. Observed a small increase in the percentage of fast type IIa fibres post-space flight 86. In this connection it would be possible to expect augmentation of increase the rate of rise of evoked twitch contraction a muscle. However, as show our data time-to-peak of a muscle after flight is increased. With these data are compounded and earlier received data in simulated conditions 46, 48, 49, 50, 51.
The rapid nature of the isometric changes, i.e., twitch duration, may be related to alterations in SR function 11, 26. The primary factor (mechanism) of the explanation for these the changes may be a reduction in the rate at which Ca2+ is dissociated from the myofibrillar proteins 9. Dissociation would occur more slowly if the rate of Ca2+ re-uptake by the SR was decreased. Such a decrease has been found following disuse 34. A reduced rate of Ca2+ dissociation from myofibrillar proteins might be expected due not only to increase the time course of the twitch response but also allow more force to be generated, since cross-bridges will continue to be formed while Ca2+ is available in the sarcoplasm. These effects on the SR would be difficult to observe as the effects on Pt would be masked by atrophy but are of interest on the assumption that the twitch changes are due to SR alterations. The changes in the kinetics the mechanical responses at paired stimulation (see Figure 2, C) might also be explained by altered development of Ca2+ kinetics in the muscle used in the experimental. At any given interpulse interval the relative increase in force of contraction after long-term space flight effect was significantly less compared to the pre-flight value.
Effects of Space Flight on Force-Velocity Relationship
The rate of rise of evoked contraction (the shape of the force-time curve) in response to electrical stimulation of the n. tibialis with a frequency of 150 impulses⋅s−1 calculated according to the relative scale showed only minor changes due to microgravity, which agrees with the observations 87 of relatively constant mechanics of tetanic contraction theory of muscle contraction 74. Similar to the flight condition (a 120 days), we have previously shown that head-down tilt no significant changes in the force-velocity characteristics of human the TS muscle 44, 47 and this observation agrees with the data obtained earlier by Witzmann et al. 87, who have shown that there were no significant changes in the force-velocity characteristics of rat soleus, extensor digitorum longus or superior, and vastus lateralis muscles after 21 days of immobilization. If the shape of the force-time curve is determined by the net rate of formation and disruption of the cross-bridges 74 which are proportional to the activity of myo-ATP-ase 75, then it may be assumed that the cycling of cross-bridges and activity of myo-ATP-ase varied slightly (or not at all) during the effects of microgravity. Therefore, one can assume that inactivity of myo-ATP-ase is reason for changing the kinetics of contraction after weightlessness. This supports the hypothesis that the changes of isometric contractile properties of the microgravity skeletal muscles being in microgravity result not from a change of contractile proteins but from any other factor/s, possible, from change in the function on SR. The increase of the normalized rate of tension development (dp/dt), that was recorded during prolonged space travel was consistent with the finding that myo-ATP-ase activity and maximal velocity of shortening have been enhanced during space flight which agrees with the observations to disuse 80. It would therefore seems reasonable to conclude that disuse, for example weightlessness or head-down tilt, in crew prolonged space travel has little effect on either cross-bridge cycling or myosin activity 17.
In addition to, it is known, the time course of force development during electrically evoked tetani did change whereas the maximal rates tetanic force development (dPec/dt) was increase (see Figure 6). The cause of the space flights-induced increase in the maximal unloaded shortening velocity is unknown. However, increases in shortening velocity as a result of microgravity can also result from geometric alterations inside muscle structures, i.e., increase in filament spacing 66 or a decrease in the pennation angle of muscle fibers 24, 85.
On the other hand, the microgravity-induced increase in the maximal unloaded shortening velocity of the TS muscle could be due to at least three possible mechanisms: i) an increase in the fast type myosin heavy chains (MHC) protein isoform content; ii) the de novo expression of a faster MHC isoform, and/or iii) an increase in the fast myosin light chains (MLC). Previous studies at both the whole muscle 14 and single-fiber 10 level have report good correlations between the MHC isoform composition and maximal unloaded shortening velocity. Hence, the 44 % increase in the maximal rates tetanic force development (dPec/dt) in the flight triceps surae muscles could be due to the corresponding increase in the relative fast type MHC protein isoform content. Also possible that some of the increase in maximal velocity may have been due to alterations in the essential MLC isoform composition of the flight muscles 86. Increased maximal shortening velocity might have occurred due to the selective loss of thin filaments. Such alterations have already been hypothesized by some authors. Riley et al. 72 shown that a 17 day space flight (STS-78) induced disproportionate loss of actin filaments in human soleus muscle. This structural change was responsible for post-flight increase in the maximal shortening velocity and for a reduced peak stiffness in the muscle 86. These alterations would be expected to have greater effect on shortening velocity than on force 86.
Effects of Space Flight on Electromechanical Delay
During many everyday activities, the ability to produce high muscle force per time unit is more important than the ability to generate high force. The rate of force development depends on many factors, in particular, on the duration of the excitation-contraction process, on the force-velocity properties of muscle fibers (even during isometric contraction due to tendon structure deformity) and SEC stiffness. Therefore, decreased MTC stiffness may be the reason for the decreased rate of development of the TS muscle contraction after unloading, since SEC stiffness in known to be an important factor determining the rate of muscle force development 9. In other words, as indicated in the Introduction, the time of SEC stretching by contractile elements determines the EMD value 38. Therefore, changes in MTC (SEC) stiffness after hyperactivity/hypoactivity mainly explain changes in EMD. In addition, all the structures of the SEC, classically composed of an active part (located in myofibrils) and a passive part (mainly aponeurosis and tendon) 88, could contribute differently to EMD. Nordez et al. 69 used the noninvasive methodology (a real-time ultrasonography) determine the relative contribution of the passive part of the SEC (47.5 ± 6.0 % of EMD) and each of the two main structures of this component (aponeurosis and tendon, representing 20.3 ± 10.7 % and 27.6 ± 11.4 % of EMD, respectively).
This study demonstrated that the rate of isometric voluntary contraction development performed after the instruction to exert the fastest contraction had decreased after unloading, which confirms the theoretical relationship between the decreased muscle MTC stiffness and decreased rate of transmission of the contractile force and therefore the rate of contraction development. This data was obtained for the first time and is indicative of the increase of MTC stiffness after PT under conditions of muscular system unloading. Our results in the present study indicate that PT during space flight results in an increase in MTS (MTC), whereas a lack of PT is associated with a decrease of this parameter 53. Therefore, PT decreases the EMD under muscular system unloading conditions.
The changes in elastic properties due to PT have been well documented. Endurance training resulted in an increase in the SEC stiffness in the soleus muscle of rats, associated with an increase in type I fibres 31. Both jump and endurance training also appear to increase both collagen concentration 23, 58 and muscle passive stiffness. The soleus rat muscles submitted to plyometric training had more fast twitch fibres and a lower SEC stiffness than controls 3, 83. Malisoux et al. 63 reported that human subjects given 8 weeks of maximal effort stretch-shortening cycle exercise training tended to have an increase in the proportion of type II fibres in their vastus lateralis muscles. Kubo et al. 60 reported that the MTS is greater in long distance runners than in untrained individuals.
The results obtained correlate well with data on the decreased EMD after isometric training under bed-rest conditions 60, 61. The EMD changes during training are mainly attributed to changes in the tendon structure, but it should be noted that tendon stiffness is always increased during any physical activity, both during endurance 12 and isometric training 60, 61. Furthermore, EMD changes correlate very closely with muscle MTC and especially with the active SEC fraction during training 22, 63.
This study demonstrated that the EMD is sensitive enough and may serve as an indirect marker for measurements of muscle MTC stiffness in order to determine the chronic adaptation of the musculo-tendinous structure of the muscle to the mechanical unloading during PT under real or simulated microgravity conditions. The data obtained demonstrates that a 180-day of actual microgravity led to decrease the TS mechanical properties and although the adverse effects were reduced thanks to PT, they were not completely prevented by the training program. This permits us to suppose that the volume of exercises and especially the intensity of exercises performed did not exceed the threshold level required for the complete prevention of changes in mechanical properties.
The mechanical muscle responses obtained and the list of physical exercises received from the study subjects, as well as from members of long-term (6-month) space missions confirm the importance of PT in preserving muscle function and the capacity to work during long-term stays under microgravity conditions. Within the framework of this study, the PT program contained mainly low intensity exercises. The inclusion of exercises with higher load and higher intensity into the training process would contribute to a more effective exercise program for the training of skeletal muscles and it would reduce the total training time under zero gravity conditions. In general, PT under microgravity conditions allows subjects to create an increased functional reserve and reduces the effect of unloading observed under real microgravity conditions.
In conclusion, the results presented here show that mechanical properties in human the TS muscle were altered after prolonged space flight. The comparison of the mechanical alterations recorded during voluntary contractions and in contractions evoked by electrical stimulation of the motor nerve (for all seven cosmonauts studied MVC produced more decreased then Po after the space flight), suggests that weightlessness not only modifies the peripheral processes associated with contractions, but also changes central and/or neural command of the contraction.
Conclusion and Recommendations
The main result of our study shows the considerable reduction of contractile functions of the TS muscle, as a muscle undergoing more changes in comparison with other muscles at unloading 2. It follows that contractile characteristics of muscle depend, among others, on background of its activity. In this connection the necessity of analysis of training process arose which shown that crewmembers of space missions used cyclic type exercises as main means of training, and it appears that this was not enough for maintaining of functional characteristics of neuromuscular apparatus as executive “work” apparatus in crewmembers of prolonged space flights. In this connection it is necessary to take into account functional possibilities of the system like neuromuscular apparatus, which suffers primarily in space flight conditions, while developing the training process.
The reduction of contractile functions in space missions crewmembers supports the idea that “protection” of skeletal muscles during long-term space flight requires physical training with more effective program. The program of physical training with the use of such means of training process as treadmill and bicycle ergometer improves aerobe capacity by providing of training of cardiovascular and respiratory systems 34, but not power characteristics of muscle apparatus. That is why the reduction of contractile functions of muscles after space flight supports the idea that “protection” of muscles during space flight requires physical training with more effective training program including in particular high-intensive and/or “explosive” exercises which increase power characteristics of muscles 73, 79, and are promising for avoiding muscles atrophy and “weakness”. Besides, analysis of training process used in space flight has not revealed specific exercises aimed at the training of plantar flexors that could be additional factor in the reduction of contractile functions of muscles.
Disclosures
The author has no conflict of interest in the study and has not received external financial support for the preparation of this manuscript
Acknowledgements
The author gratefully acknowledges the crewmembers of the Space Station «MIR» who graciously gave their time and energy to this research. Without their efforts this project would have been impossible. He thanks the medical staff of Yuri A. Gagarin Cosmonauts Training Center at Star City, Moscow, V. Morgun, M.D., Ph.D., D.Sci., V. Pochuev, M.D., Ph.D., A. Polyakov, M.D., Ph.D., E. Kobzev, M.D., Ph.D., A. Kulev, M.D., and V. Dronov, engineer, for their contributions in organizing the study and the numerous unnamed people who assisted our research.
This work was supported by the Fonds SSC -Institute of Biomedical Problems of the Russian Academy of Sciences.
References
- 1.Adams G R, Hather B M, Dudley G A. (1994) Effect of short-term unweighting on human skeletal muscle strength and size. Aviat Space Environ Med. 65, 1116-1121.
- 2.Akima H, Kubo K, Imai M, Kanehi H, Suzuki Y et al. (2001) Inactivity and muscle: effect of resistance training during bed rest on muscle size in the lower limb. Acta Physiol Scand. 172-269.
- 3.Almeida-Silveira M I, Pérot C, Pousson M, Goubel F. (1994) Effects of stretch-shortening cycle training on mechanical properties and fibre type transition in the rat soleus muscle. Pflüg Arch. 427-289.
- 4.Antonutto G, Bodem F, Zamparo P, di Prampero PE. (1998) Maximal power and EMG of lower limbs after 21 days space flights in one astronaut. , J Gravit Physiol 5, 63-66.
- 5.Antonutto G, Capelli C, Cirardis M, Zamparo P. (1999) di Prampero PE. Effects of microgravity on maximal power of lower limbs during very short efforts in humans. , J Appl Physiol 86, 85-92.
- 6.Steven B R, Guilliams M E, Greenisen M C. (1997) Resistance exercise prevents plantar flexor deconditioning during bed rest. Aviat Space Environ Med. 29, 1462-1468.
- 7.Barany M. (1967) ATPase activity of myosin correlated with speed of muscle shortening. , J Genet Physiol 50, 197-216.
- 8.Blazevich A J, Horne S, Cannavan D, Coleman D R, Aagaard P. (2008) Effect of contraction mode of slow-speed resistance training on the maximum rate of force development in the human quadriceps. Muscle Nerve. 38, 1133-1146.
- 9.Bojsen-Møller J, Magnusson S P, Rasmussen L R, Kjaer M, Aagaard P. (2005) Muscle performance during maximal isometric and dynamic contractions is influenced by the stiffness of the tendinous structures. , J Appl Physiol 99, 986-994.
- 10.Bottinell R, Betto R, Schiaffino S, Reggiani C. (1994) Unloaded shortening velocity and myosin heavy chain and alkali light chain isoform composition in rat skeletal muscle fibers. , J Physiol 478-341.
- 11.Solard R J. (1977) Relative capabilities of sarcoplasmic reticulum in fast and slow mammalian skeletal muscle. , J Physiol 266-587.
- 12.Buchanan C I, Marsh R L. (2001) Effects of long-term exercise on the biomechanical properties of the Achilles tendon of guinea fowl. , J. Appl. Physiol 90, 164-171.
- 13.Biggs N L, Blanton P L, Lehr R R. (1973) Electromyographic investigation of the relative activity among four components of the triceps surae. , Amer J Phys Med 52, 30-41.
- 14.Caiozzo V J, Herrick R E, Baldwin K M. (1992) Response of slow and fast muscle to hypothyroidism: maximal shortening velocity and myosin isoforms. , Amer J Physiol 263-86.
- 15.Cavanagh P R, Komi P V. (1979) Electromechanical delay in human skeletal muscle under concentric and eccentric contractions. , Eur J Appl Physiol 42, 159-163.
- 16.Clément G, Gurfinkel V S, Lestienne F. (1985) Mechanisms of posture maintenance in weightlessness. Vestibular and Visual Control on Posture and Locomotor Equilibrum, ed. Black, I , Basel, Switzerland: Karger 158-163.
- 18.Convertino V A. (1990) Physiological adaptations to weightlessness: effects on exercise and work performance. Exerc and Sport Sci Rev (Pandolf KB, Holloszy JO eds.) , Baltimore, MD: Williams 119-166.
- 19.Darr K C, Schultz E. (1989) Hindlimb suspension suppresses muscle growth and satellite cell proliferation. , J Appl Physiol 67, 1827-1834.
- 20.Di Fabio RP. (1987) Reliability of computerized surface electromyography for determining the onset of muscle activity. Phys Ther. 67, 43-48.
- 21.Duchateau J, Hainaut K. (1990) Effects of immobilization on contractile properties, recruiment and firing rates of human motor units. , J Physiol 422-55.
- 22.Ducomps C, Mauriege P, Darche B, Combes S, Lebas F et al. (2003) Effects of jump training on passive mechanical stress and stiffness in rabbit skeletal muscle: role of collagen. Acta Physiol Scand. 178-215.
- 23.Edgerton V R, Barnard R J, Peter J, Meier A. (1975) Properties of immobilized hind limb muscles of the galago senegalensis. , Exp Neurol 46, 115-131.
- 24.Eremin A I, Bazhanov Marishchuk VL, Stepantsov Dzhamgarov TT. (1969) Human training in conditions of long-term hypokinesia. Prob of Space Biol. , Moscow: Nauka 191-199.
- 25.25 Fitts RH, Riley D R, Widrick J J. (2000) Physiology of a microgravity environment. Invited Review: Microgravity and skeletal muscle. , J of Appl Physiol 89, 823-839.
- 26.Contractile biochemical. (1980) and histochemical properties of thyrotoxic rat soleus muscle. , Am J of Physiol 238-15.
- 27.Friedrich J A, Brand R A. (1990) Muscle fiber architecture in the human lower limb. , J of Biomech 23, 91-95.
- 28.Gabriel D A, Basford J R. (2001) An K-N Training-related changes in the maximal rate or torque development and EMG activity. , J Electromyog Kines 11, 123-129.
- 29.Gardetto P R, Schluter J M, Fitts R H. (1989) Contractile function of single muscle fibers after hindlimb suspension. , J of Appl Physiol 66, 2739-2749.
- 30.Goldspink G, Tabary C, Tabary J C, Tardien C, Tardien G. (1974) Effect of denervation on the adaptation of sarcomere length of the muscle. , J of Physiol 236-733.
- 31.Goubel F, Marini J F. (1987) Fibre type transition and stiffness modification of soleus muscle of trained rats. Pflüg Arch. 410-321.
- 32.Greenleaf J E, Bulbulian R, Bernauer E M, Haskell W L, Moore T. (1989) Exercise-training protocols for astronauts in microgravity. , J of Appl Physiol 67, 2191-2204.
- 33.Grigor'ev A I, Dietlein L F, Kozlovskaya I B, Sawin C F. (2001) Preventive measures in short- and long-term space flights. Kosm Biol Med Health Work Capacity, and Safety of Space Crews. Joint US/Russian publication (Pestov ID, Dietlein LF, eds.) , Moscow: 4, 252-309.
- 34.Grigor'eyva L S, Kozlovskaya I B. (1987) Effect on weightlessness and hipokinesia on velocity and strength properties of human muscle. Kosm Biol and Aviakosm Med. 21, 27-30.
- 35.Grosset J-F, Julien P, Lambertz D, Ch Pérot. (2009) Paired changes in electromechanical delay and musculo-tendinous stiffness after endurance or plyometric training. , Eur J Appl Physiol 105-131.
- 36.Häkkinen K, Keskinen K L. (1989) Muscle cross-sectional area and voluntary force production characteristics in elite strength- and endurance-trained athletes and sprinters. , Eur J of Appl Physiol 59, 215-220.
- 37.Häkkinen К, Viitasalo J T, Komi P V. (1980) Die Workung unterschiedlich kombinierter konzentrischer und exzentrischer Muskelarbeit auf Kraft-Zeit-Merkmale der Beinstreckmuskulatur. Leistungssport. 10, 374-381.
- 38.Hill A V. (1938) The heat of shortening and the dynamic constants of muscle. Proc Royal Soc Lond Ser B Containing Papers of a Biol. Character. 126-136.
- 39.Johnson M A, Polgar J, Weightman D, Appleton D. (1973) Data on the distribution of fibre types in thirty-sex human muscles: an autopsy study. , J of Neurol Sci 18, 111-129.
- 40.Kawakami Y, Abe T, Fukunaga T. (1993) Muscle-fiber pennation angles are greater in hypertrophied than in normal muscles. , J of Appl Physiol 74, 2740-2744.
- 41.Kawakami Y, Akima H, Kubo K, Muraoka Y, Hasegawa H et al. (2001) Changes in muscle size, architecturem and neural activation after 20 days of bed rest with and without resistance exercise. , Eur J of Appl Physiol 84, 7-12.
- 42.Kim D H, Witzmann F A, Fitts R H. (1982) Effects of disuse on sarcoplasmic reticulum in fast and slow skeletal muscle. , Am J of Physiol 243-156.
- 43.Yu Koryak. (1994) A comparison of contractile properties in human leg muscles. , Physiol J 40, 30-38.
- 44.Yu Koryak. (1995) Contractile properties of the human triceps surae muscle during simulated weightlessness. , Eur J Appl Physiol 70, 344-350.
- 45.Yu Koryak. (1995) Skeletal muscle responses to lower limb "dry" immersion in human. Aviat Space and Environ Med. 66-473.
- 46.Yu Koryak. (1996) Changes in the action potential and contractile properties of skeletal muscle in human's with repetitive stimulation after long-term dry immersion. , Eur J of Appl Physiol 74, 496-503.
- 47.Yu Koryak. (1998) Influences of 120-days 6 degree head-down tilt bed rest on the functional properties of neuromuscular system in man. Aviat Space and Environ Med. 69, 766-770.
- 48.Yu Koryak. (1998) Electromyographic study of the contractile and electrical properties of the human triceps surae muscle in a simulated microgravity environment. , J of Physiol 510-287.
- 49.Yu Koryak. (1998) Effect of 120 days of bed-rest with and without countermeasures on the mechanical properties of the triceps surae muscle in young women. , Eur J of Appl Physiol 78, 128-135.
- 50.Yu Koryak. (1999) Effects of long-term simulated microgravity on neuromuscular performance in men and women. , Eur J of Appl Physiol 79, 168-175.
- 51.Yu Koryak. (2000) The effects of long-term muscle unloading on the contractile and electrical properties of the human triceps surae. Proc 13th Man in Space: The Future of Human in Space, Symposium , Greece 236-237.
- 52.YuA Koryak. (2011) Adaptation of Human Skeletal Muscle to the Load Change. Experimental study Germany: LAP.
- 53.Koryak Yu A. (2012) Contractile properties and musculo-tendinous stiffness of the triceps surae muscle and their changes as a result of continuous bed rest. , Physiol J 58, 66-79.
- 54.Koryak Yu A. (2014) Influence simulated microgravity on mechanical properties in the human triceps surae muscle in vivo. I. Effect of 120-days of bed-rest without physical training on human muscle musculo-tendinous stiffness and contractile properties in young women. , Eur J Appl Physiol 114-1025.
- 55.Yu Koryak. (1997) Kozlovskaya I. The effects of a 115-day space flights on neuromuscular function in crewman. Proc 12th Man in Space: The Future of Humans in Space. Simposium , Washington, DC 231.
- 56.Yu Koryak. (2000) Kozlovskaya IB. Changes in electrically evoked and voluntary skeletal muscle contractions during long-term space flights. IXth Inter Symp "Motor Control" (Kossev A. , ed), Bulgaria 6.
- 57.Yu Koryak, Siconolfi S F, Kozlovskaya I B, Gilbert J H, Layne C S. (1997) Maximal voluntary (MVC), tetanic (Po) and single twitch (Pt) contractions before & after space flight. , FASEB J 1408.
- 58.Kovanen V, Suominen H, Heikkinen E. (1980) Connective tissue of ‘‘fast’’ and ‘‘slow’’ skeletal muscle in rats — effects of endurance training. , Acta Physiol. Scand 108-173.
- 59.YaM Kots, Absalyamov T M, Zorin V P, YuA Koryak, Kuznetsov S P et al. (1976) A modification of tendometric method of measuring contraction force of single muscles in humans. , Fiziol. Cheloveka 2, 1045-1048.
- 60.Kubo K, Akima H, Kouzaki M, Ito M, Kawakami Y et al. (2000) Changes in the elastic properties of tendon structures following 20 days bed-rest in humans. , Eur J of Appl Physiol 83, 463-468.
- 61.Kubo K, Kanehisa H, Kawakami Y, Fukunaga T. (2000) Elastic properties of muscle–tendon complex in long-distance runners. , Eur. J. Appl. Physiol 81, 181-187.
- 62.Lambertz D, Pérot C, Kaspranski R, Goubel F. (2001) Effects of long-term space flights on mechanical properties of muscles in humans. , J of Appl Physiol 90, 179-188.
- 63.Malisoux L, Francaux M, Nielens H, Theisen D. (2006) Stretch-shortening cycle exercises: an effective training paradigm to enhance power output of human single muscle fibers. , J. Appl. Physiol 100-771.
- 64.Mayer R F, Burke R E, Toop J, Hodgson J A, Kanda K et al. (1981) The effect of long-term immobilization on the motor unit population of the cat medial gastrocnemius muscle. Neurosci. 6, 725-739.
- 65.Muraoka T, Muramatsu T, Fukunaga T, Kanehisa H. (2004) Influence of tendon slack on electromechanical delay in the human medial gastrocnemius in vivo. , J Appl Physiol 96, 540-544.
- 66.Murray M P, Gutten G H, Baldwin J M, Gardner G M. (1976) A comparison of plantor flexion torque with and without the triceps surae. Acta Orthopaed Scand. 47, 122-124.
- 67.Orizio C, Gobbo M, Veicsteinas A, Baratta R V, Zhou B H et al. (2003) Transients of the force and surface mechanomyogram during cat gastrocnemius tetanic stimulation. , Eur J Appl Physiol 88, 601-606.
- 68.Narici M V, Kayse B, Barattini P, Cerretelli P. (1997) Changes in electrically evoked skeletal muscle contractions during 17-day space flights and bed rest. , Inter J Sports Med 18, 290-292.
- 69.Nordez A, Gallot T, Catheline S, Guevel A, Cornu C et al. (2009) Electromechanical delay revisited using very high frame rate ultrasound. , J. Appl. Physiol 106-1970.
- 70.Powell P, Roy R R, Kanim P, Bello M A, Edgerton V R. (1984) Predictability of skeletal muscle tension from architectural determinations in guinea pig hindlimbs. , J of Appl Physiol 57, 1715-1721.
- 71.Recktenwald M R, Hodgson J A, Roy R R, Riazanski S, McCall G E et al. (1999) Effect of space flights on rhesus quadrupedal locomotion after return to 1G. , J of Neurophysiol 81, 2451-2463.
- 72.Riley D A, JLW Bain, Thopson J L, Fitts R H, Widrick J J et al. (2000) Decreases thin filament density and length in human atrophic soleus muscle fibers after spacefligth. , J of Appl Physiol 88, 567-572.
- 73.ale D G, Mc Comas AJ, Mac Dougall JD, Upton A R. (1982) Neuromuscular adaptation in human thenar muscles following strength and immobilization. , J of Appl Physiol 53, 419-424.
- 74.Simmons R M, Jewell B R. (1974) Mechanics and models of muscular contraction. Res Adv Physiol. 9, 87-147.
- 75.St-Pierre D, Gardiner P F. (1985) Effect of "disuse" on mammalian fast-twitch muscle: joint fixation compared with neurally applied to tetrodotoxin. Exp Neurol. 90, 635-651.
- 76.Stepantsov Tikhonov MA, Eremin A V. (1972) Physical training as a method to prevent hypodynamia syndrome. , Kosm. Biol. Aviakosm. Med 6, 64-69.
- 77.Steven L, Mounier Y. (1992) Ca2+ movements in sarcoplasmic reticulum of rat soleus fibers after hindlimb suspension. , J of Appl Physiol 72, 1735-1740.
- 78.Steven L, Mounier Y, Holy X, Falempin M. (1990) Contractile properties of rat soleus muscle after 15 days of hindlimb suspension. , J of Appl Physiol 68, 334-340.
- 79.Trappe S, David Costill D, Ph Gallagher, Creer A, Peters J R et al. (2009) Exercise in space: human skeletal muscle after 6 months aboard the International Space Station. , J Appl Physiol 106, 1159-1168.
- 80.Unsworht B R, Witzmann F A, Fitts R H. (1982) A comparison of rat myosine from fast and slow skeletal muscle and the effect of disuse. , J of Biol Chem 25, 15127-15136.
- 81.M Van Cutsem, Dechateau J. (1998) Hainaut K Changes in single motor unit behaviour contribute to the increase in contraction speed after dynamic training in humans. , J of Physiol 513-295.
- 82.Vandervoort A A. (1983) Mc Comas AJ. A comparison of the contractile properties of the human gastrocnemius and soleus muscles. , Eur J of Appl Physiol 52, 435-440.
- 83.Watt P W, Kelly F J, Goldspink D F, Goldspink G. (1982) Exercise induced morphological and biochemical changes in skeletal muscles of the rat. , J. of Appl. Physiol 53, 1144-1151.
- 84.Weiss A D. (1965) The locus of reaction time change with set, motivation and age. , J Gerontol 20, 60-64.
- 85.Wickiewicz T L, Roy R R, Powell P L, Edgerton V R. (1983) Muscle architecture of the human lower limb. Clin Orthopaedics. 179-275.
- 86.Widrick J J, Knuth S T, Norenberg K M, Romatowski J C, JLW Bain et al. (1999) Effect of a 17 day space flights on contractile properties of human soleus muscle fibres. , J of Physiol 516-915.
- 87.Witzmann F A, Kim D H, Fitts R H. (1982) Recovery time course in contractile function on fast and slow skeletal muscle after hind limb immobilization. , J of Appl Physiol 52, 677-682.
Cited by (1)
- 1.Koryak Yu. A., Kukoba T. B., Babich D. R., 2023, Changes in the Functions and Architecture of Human Skeletal Muscles during 21-Day Unloading of the Locomotor System without Physical Exercise, Human Physiology, 49(2), 115, 10.1134/S036211972260045X